Sauropterygia: Pliosauroidea: Rhomaleosauridae
Taxa on This Page
- Leptocleidus X
- Rhomaleosauridae X
- Rhomaleosaurus X
- Simolestes X
- Yuzhoupliosaurus X
The Rhomaleosaurs

The Rhomaleosauridae were the earliest of the giant sauropterygian predators, rivaling the biggest ichthyosaurs in size. They are also relatively primitive pliosauroids. They differ from conventional pliosaurs in their longer neck and crocodile-like head. For some thirty or forty million years they were the top predators of the ocean, attaining 5 to 7 metres in length. Genera may include Rhomaleosaurus (=Thaumatosaurus), Yuzhoupliosaurus, Macroplata, Sthenarosaurus, and Simolestes, but it is not certain if any or all of these genera are actually related, or whether they are the result of evolutionary convergence due to similar lifestyle. Some species have previously been identified as species of Plesiosaurus. In certain respects the rhomaleosaurs seem to be intermediate between the plesiosauroid and pliosauroid lineages. The group may also include Leptocleidus. However, the phylogenetic relationships of Early Jurassic Sauropterygia are poorly known, and the whole group is in need of study and revision.
In an email to one of us (MAK) on this subject (24 Aug 1999) Darren Naish says:
As you know, plesiosaur affinities are a mess, and no one can say what the true picture was at present. I do suspect that rhomaleosaurs are primitive pliosauroids, and that pliosauroids are primitive plesiosaurs (i.e. they diverged before things like Plesiosaurus, cryptoclidids and elasmosaurids), but I am not convinced that rhomaleosaurs are close to true pliosaurids like the Liopleurodon group and Peloneustes. However, I also believe that Leptocleidines are highly specialized rhomaleosaurs - both groups share the same pattern of premaxillary foramina and both have a peculiar trough on the inside of the lower jaw. This is exciting as it means that rhomaleosaurs lived on as miniaturized predators that invaded estuarine/
??freshwater environments.
 Image: Right; Rhomaleosaurus. Early Jurassic. Length 3.5 to 6 meters. Illustration by John Sibbick - in David Norman, Prehistoric Life: the Rise of Vertebrates. Left: Leptocleidus from Brian Choo, donated to Classroom Clipart for educational use. MAK991206.
Image: Right; Rhomaleosaurus. Early Jurassic. Length 3.5 to 6 meters. Illustration by John Sibbick - in David Norman, Prehistoric Life: the Rise of Vertebrates. Left: Leptocleidus from Brian Choo, donated to Classroom Clipart for educational use. MAK991206.
Rhomaleosaurus
This important genus includes a number of species, some of which grew quite large. These were powerfully built pliosaurs, with long toothy jaws which hunted both by site and smell and tore their prey apart with jerks of the head, much as a modern crocodile does. Think of a crocodile crossed with a modern killer whale.
Cruickshank et al. (1991), showed that the skull of this predator is designed in a way that the animal could swim with its mouth slightly open, enabling water to pass through scoop-shaped openings in the roof of the mouth, through a channel where the smell sensors were located (Jacobson's organ), then out external nostrils on the snout. This arrangement -- the reverse of what might be expected in a normal air-breathing animal, would have enabled Rhomaleosaurus to hunt its prey by smell, just sharks do.
The following material is taken from their short article in Nature, and shows how the skull of this marine carnivore was designed to allow water to flow in through the nostrils (nares), thus enabling it to smell the water:
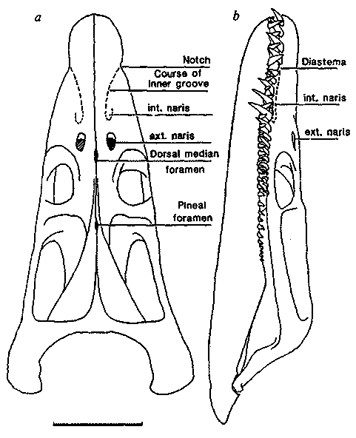 The dorsally placed external nostrils of plesiosaurs are usually regarded as an adaptation to breathing in those extinct marine reptiles. We suggest instead that the narial system was used in underwater olfaction. The internal nares are anterior to the external nares. Hydrodynamic pressure during swimming forced water into the mouth, along palatal grooves into the scoop-shaped internal nares and up short ducts, presumably lined with olfactory epithelia. Alternatively, or additionally, the so far unlocated Jacobson's organ detected particulate matter. The water was sucked out through the external nares by hydrodynamic pressure generated by fast flow over the convex upper surface of the head.
The dorsally placed external nostrils of plesiosaurs are usually regarded as an adaptation to breathing in those extinct marine reptiles. We suggest instead that the narial system was used in underwater olfaction. The internal nares are anterior to the external nares. Hydrodynamic pressure during swimming forced water into the mouth, along palatal grooves into the scoop-shaped internal nares and up short ducts, presumably lined with olfactory epithelia. Alternatively, or additionally, the so far unlocated Jacobson's organ detected particulate matter. The water was sucked out through the external nares by hydrodynamic pressure generated by fast flow over the convex upper surface of the head.
The internal nares strongly resembles the NACA Duct (National Advisory Committee for Aeronautics) design of air scoops on aircraft, whose opening is flush with the surface. This design deepens and widens in the direction of fluid flow and ends in a transverse scoop, suggesting that the internal nares acted as a ram scoop for water flowing backwards along the inner groove....
Were the nares used for respiration? Both internal and external nares would have been partly occluded by soft tissue and any measurement of the bony outlines will only give a maximum dimension. In the very similar R. zetlandicus, sculpture on the anterior external narial wall may indicate insertions for muscles and ligaments controlling a narial valve. In R. megacephalus each internal nares had a functionally clear area of 2.0 cm2 or less, which seems disproportionately small for a 5-meter long Rhomaleosaurus. Also, the placement of the internal nares anterior to the external nares would have hindered respiratory air flow. We believe that Rhomaleosaurus breathed through its open mouth, contrary to the conventional view that all respiration occurs through the narial system. Water would have been excluded from the trachea during normal swimming and in heavy seas, by a glottal valve and a large muscular tongue. The reconstruction of anteriorly hinged nasal flaps on the external nares would presume automatic closure if inspiration were attempted through them. We therefore believe that the dorsal placement of the external nares is thus irrelevant for respiration, unlike in crocodilians and cetaceans, which have full secondary palates. In these two groups with secondary palates, respiration is associated with airborne olfaction, but in the case of this pliosaur we believe the adaptation to be for the sampling of water and not air.
 Rhomaleosauridae:
Rhomaleosauridae:
Range: Early Jurassic to Middle Jurassic (middle Cretaceous?).
Phylogeny: Pliosauroidea::: Pliosauridae + *: Yuzhoupliosaurus + (Rhomaleosaurus
+ (Simolestes + Leptocleidus)).
Characters: skull relatively large (12-17% total length); rostrum short-moderate length; rostrum distally expanded and spatulate; some large, caniniform teeth (perhaps associated with croc-like bight, twist & tear tactics); neck short (20-30% body length); cervical centra short & compact.
Note: [1] precisely because of the common requirements of a croc-style hunter, these features may be the result of convergence. [2] Since Yuzhoupliosaurus is known only in small part, the reconstruction shown here should be taken as that of a fair average sort of rhomaleosaur.
Links: The Plesiosaur Site - Species; Rhomaleosauridae after O'Keefe, 2001; Plesiosauria Translation and Pronunciation Guide. ATW020703.
 Yuzhoupliosaurus: Y. chengjiangensis Zhang 1985.
Yuzhoupliosaurus: Y. chengjiangensis Zhang 1985.
Range: Middle Jurassic of China
Characters: ~4m?; lower jaw containing 5 pairs of large teeth; about 23 or 24 smaller teeth; cervical vertebrae high and short; anterior cervical ribs double-headed; posterior cervical ribs single-headed (a more derived form); scapula triradiate; coracoids elongated anteroposteriorly; clavicles well developed; believed to be freshwater form.
Phylogeny: Rhomaleosauridae: (Rhomaleosaurus + (Simolestes + Leptocleidus)) + *.
Note: known only from lower jaw, vertebrae & fragments of pectoral girdle.
Links: Plesiosauria Translation and Pronunciation Guide.
References: Zhang 1985)
 Rhomaleosaurus: Seeley 1874.
Rhomaleosaurus: Seeley 1874.
Range: Early Jurassic (Hettangian to Toarcian)
Phylogeny: Rhomaleosauridae:: Simolestes + Leptocleidus) + *.
Characters: neck short, with ~15. vertebrae; neural arches bases of (cervical? all?) vertebrae extending along entire length of centrum; ~25 dorsals; 4-5 sacrals; 2 pairs sacral ribs fused to ilium; humerus larger than femur.
Rhomaleosaurus megacephalus Stutchbury 1846: Hettangian of Somerset, England. This species represents the earliest species of Rhomaleosaurus. It averaged about 5 meters in length. The long crocodile-like skull alone was about 90 cm. This species was previously described as a species of Eurycleidus.
Rhomaleosaurus propinquus Blake 1876: from the Toarcian of Whitby, England, is similar in size and build to R. victor (below). It is characterized by a distinctive lower jaw.
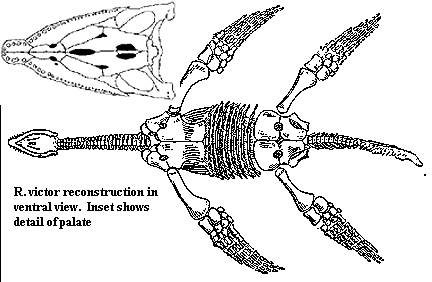
Rhomaleosaurus victor Fraas 1910a: from the Toarcian of Germany and England, was 3 to 4 metres in length. This "gracile" species has also been identified as Thaumatosaurus ("miraculous lizard,") and Eurycleidus. This long-necked form is similar in proportions to Plesiosaurus, but with a somewhat shorter neck. It is distinguished from other rhomaleosaurs by its proportionately much larger paddles, smaller and broader head, and relatively longer neck. MAK991206 Update cladistic analysis shows Rhomaleosaurus to be a paraphyletic assemblage consisting of a number of distinct if closely related forms and sevral different lineages (Smith & Dyke 2008). In keeping with the current trend in phylogenetics this means that many of these taxa will have to be renamed [1]. It has since been given its own genus, Meyerasaurus (Smith & Vincent 2010); unfortunately the superb original name Thaumatosaurus ("miraculous reptile"), familar to generations of scientists and palaeo geeks (including yours truely) was an invalid taxon and could not be reinstated. MAK111031
Rhomaleosaurus zetlandicus Phillips 1854 is known from fossil remains the Toarcian of the Yorkshire coast (Whitby) England. This was a large predator, and the largest and heaviest known species of the Rhomaleosauridae. The holotype is an incomplete skeleton about 5.3 meters long. The head makes up 74 cm (14%) and the neck 1.3 meters (25%).
Rhomaleosaurus cramptoni Carte & Bailey 1863 and R. thorntoni Andrews, also both known from the Toarcian of England - are similar in size to the contemporary R. zetlandicus, reaching 5 to 7 meters. They are now considered junior synonyms of (the same species as) R. zetlandicus.
Links: Fossils of the Lias (Incredible photo near bottom of page); Plesiosauria Translation and Pronunciation Guide; The Plesiosaur Site - Imagese (beautiful photo of R. victor); The Plesiosaur Site - Species; The Plesiosaur Site - Home Page; ?????? ??? (Japanese); JURÁSSICO Portuguese); wm851s.htm detailed sketches of R. propinquus).
References: Taylor 1992). MAK991206, ATW020704
Simolestes: Andrews 1909. S. indicus Bardet et al. 1991 (= Plesiosaurus indicus Lydekker 1877), S. keileni Godefroit 1994, S. vorax Andrews 1909.
Range: Early Jurassic of Europe & India (not Africa -- this turned out to be a fragment from a teleosaurid). S. vorax is from the Callovian, Oxford Clay, Peterborough, England.
Phylogeny: Rhomaleosauridae:::
Leptocleidus + *.
Characters: skull short & broad; rostrum not elongated; "rosette shaped symphysis" at end of lower jaw (probably an artifact) with 5-6 (pairs?) caniniform teeth; cervical vertebral centra (~20) with diameter twice length; 30-32 dorsals; 18+ caudals.
Links: NEW PLIOSAURIDS A-PLENTY; Plesiosauria Translation and Pronunciation Guide; Color Banding in Dinosaur Teeth - Reason? + plesiosaur repro.-Replies. ATW020704.
 Leptocleidus: Andrews 1922. L. capensis Cruickshank 1997 (= Plesiosaurus capensis Andrews 1911), L. clemai Cruickshank & Long 1997, L. superstes Andrews 1922.
Leptocleidus: Andrews 1922. L. capensis Cruickshank 1997 (= Plesiosaurus capensis Andrews 1911), L. clemai Cruickshank & Long 1997, L. superstes Andrews 1922.
Range: Early Cretaceous to middle Cretaceous of Europe, South Africa & Australia
Phylogeny: Rhomaleosauridae::
Simolestes + *.
Characters: small-bodied (2-3 m); skull triangular; crest running from nasals posteriorly and merging with prominent sagittal crest; forwardly pointing expansion ('cockscomb') on the squamosal mid-line, at rear of parasagital crest, very similar to that seen in Late Cretaceous Polycotylidae; crest flanked by deep grooves or depressions which cause orbital rims to stand up from the general profile of the skull; constricted maxilla- premaxilla junction; spatulate mandibular symphysis; dorsomedially directed trough on prearticular & adjacent bones of lower jaw, similar  to Rhomaleosaurus; 21 pairs of teeth in upper jaw (5 premaxillary + 16 maxillary), ~35 in lower; no vertebrae compressed; 13-24 cervical vertebrae; cervical ribs single-headed; cervical centra with deep central depression; scapulae small & widely separated; "thin clavicular arch" along the edge of the front limb girdle, composed of two "rather large" clavicles and a "transversely elongated" interclavicle (Plesiosauria Translation and Pronunciation Guide); humerus relatively straight, with symmetrical, fan-shaped distal end; freshwater or estuarine habit.
to Rhomaleosaurus; 21 pairs of teeth in upper jaw (5 premaxillary + 16 maxillary), ~35 in lower; no vertebrae compressed; 13-24 cervical vertebrae; cervical ribs single-headed; cervical centra with deep central depression; scapulae small & widely separated; "thin clavicular arch" along the edge of the front limb girdle, composed of two "rather large" clavicles and a "transversely elongated" interclavicle (Plesiosauria Translation and Pronunciation Guide); humerus relatively straight, with symmetrical, fan-shaped distal end; freshwater or estuarine habit.
Images: Right: opalizied remains from an Australian juvenile specimen at Fishing For Plesiosaur. Left: "Leptocleidus-type" cervical vertebra from The Plesiosaur Site.
Links: Plesiosauria Translation and Pronunciation Guide; Marine Reptiles (NOT Dinosaurs); NEW PLIOSAURIDS A-PLENTY; Crag's Lost Kingdom (?!); Image -- The Dinosauricon; Post forum2 3.pdf; Fishing For Plesiosaur; DinoWight- Leptocleidus, an Isle of Wight pliosaur; The Plesiosaur Site - Species (Best on the Web); evolution beautifully preserved undescribed specimen from Canada); Leptocleidus.htm Japanese); Eric, the pliosaur. ATW020704.
Notes:
[1] The present author (MAK) does not see the need for Linnaean binomials including generic names to conform to cladistic nomenclature. Given that these are two distinct systems, there should be nothing wrong with retaining genera as paraphyletic "grades"; however this sentiment is not shared by the majrority of systematists
Using this material. All material by ATW is public domain and may be freely used in any way (also any material jointly written by ATW and MAK). All material by MAK is licensed Creative Commons Attribution License Version 3.0, and may be freely used provided acknowedgement is given. All Wikipedia material is either Gnu Open Source or Creative Commons (see original Wikipedia page for details). Other graphics are copyright their respective owners
