
| Ornithischia | ||
| The Vertebrates | Thyreophora |
| Vertebrates Home | Vertebrate | Vertebrate |
|
Abbreviated Dendrogram
Dinosauromorpha
├─┬─Sauropodomorpha
│ └─Theropoda
│
└─Ornithischia
╞═Fabrosauridae
└─Genasauria
├─Thyreophora
│ ├─Scutellosaurus
│ └─┬─Emausaurus
│ └─Eurypoda
│ ├─Stegosauria
│ └─Ankylosauromorpha
│ ├─Scelidosauridae
│ └─Ankylosauria
└─Cerapoda
├─Ornithopoda
│ ╞═Hypsilophodontidae
│ └─Hadrosauridae
└─┬─Heterodontosauridae
└─Marginocephalia
├─Pachycephalosauria
└─Ceratopsia
|
Contents
Overview |

These are small to large specialized quadrupedal armoured forms - Early Jurassic to Late Cretaceous - length one to ten metres. There are three traditional subgroups: the primitive ancestral scelidosaurs, the unusual stegosaurs, and the heavily armoured ankylosaurs. It has been suggested that the Stegosaurs are actually more closely related to the ornithopods, but generally the following three lineages are grouped together, with the scelidosaurs as the basal (paraphyletic) grade from which the other two clades developed.
Writing about popular dinosaurs is a daunting proposition. So many have written so much and so well that little remains but to be repetitious or wrong. Given the choice between ennui and error, we will choose a little of both by discussing choices -- lineage splitting -- and how these dinosaurs made them.
The armored dinosaurs are an interesting opportunity to study radiations. The earliest dinosaurs all looked rather alike: barrel-bodied, with long necks, long, smallish heads like truncated cones, powerful hind limbs and tails, and surprisingly agile-looking forelimbs with prodigious claws. Examples of this general body plan include Herrerasaurus, Saturnalia, Thescelosaurus, and even Scelidosaurus. These forms represent all of the major groups of the first, Triassic, dinosaurian radiation: Sauropodomorpha, Theropoda, Cerapoda, and Thyreophora, respectively. These transitions can be readily understood -- if at the cost of considerable oversimplification -- as diversification into large-bodied guilds of carnivores, and of high, medium, and low browsers, respectively. It appears that either the dinosaurs possessed overwhelming selective advantages, or they were radiating into an ecological vacuum, since they managed to dominate all of these guilds by the earliest Jurassic without greatly elaborating on the basic dinosaur body plan.
At that point, however, the dynamics of dinosaur evolution began to change. Perhaps this occurred because the only serious competition left was from other dinosaurs. In any case, by the Middle Jurassic, the dinosaurs evolved many new and peculiar body forms which differed radically from the original dinosaur bauplan. These included the big-headed, bipedal tetanurans, the classic long-necked sauropods, the posturally enigmatic, short-bodied, stiff-tailed iguanodonts and their early hadrosaur and marginocephalian relatives, the heavily armored eurypods ankylosaurs and stegosaurs), and perhaps minor forms we know nothing about as yet. The Jurassic was a poor time for fossils and we are still missing large parts of that story.
The eurypod part of the tale is the part that concerns us. Here, two lineages split: the stegosaurs and the ankylosaurs. Both were elephantine, armored, low-level browsers with wide bodies presumably accommodating a powerful digestive fermentation system. Both groups are relatively uncommon dinosaurs, but both are cosmopolitan in distribution. Unlike the original radiation, these forms do not appear to have split along the fault lines of easily identified ecological guilds. The question (which it has taken us rather long to get to) is "why?" Scelidosaurus suggests that the first thyreophorans looked more or less like every other early dinosaur, with the addition of some osteoscute armor and a wedge-shaped skull. It is understandable that some lineage would diverge into armored, elephantine herbivory. But why two such bizarre and divergent forms?
The question is complicated by the fact that stegosaurs and ankylosaurs flourished at different times. Stegosaurs are known from the Cretaceous, but are primarily creatures of the Jurassic. The opposite is true of the ankylosaurs. However, there is little doubt that the split occurred in the early Jurassic. The ankylosaurs were simply less successful. The possible reasons for this may fall into one or more of the following categories: (1) some environmental condition(s) disfavored the ankylosaurian form, (2) ankylosaurs did not evolve their typical mid-Cretaceous body form for a very long time, or (3) ankylosaurs were marginalized by competition from the stegosaurs.
Reason (1) seems unlikely. Absent conditions amounting to a general mass extinction, it would be hard to imagine a case which would encourage the diversification of stegosaurs, while suppressing the ankylosaurs worldwide, for 100My or so. Condition (2) can also be eliminated. We know almost nothing about Jurassic, or even Early Cretaceous, ankylosaurs. However, it takes only one exception to disprove this hypothesis, and at least two exceptions appear to exist. Dracopelta and Tianchiasaurus are well-enough known to state that both definitive nodosaurids and ankylosaurids, respectively, existed in widely divergent locations during the Jurassic. Therefore, we can conclude that the most important condition limiting the success of the ankylosaurs was most probably competition from their cousins and fellow low-browsers, the stegosaurs.
Based on this somewhat attenuated chain of reasoning, as well as the earlier history of the thyreophoran radiation, we can tentatively identify three stages of a major radiation (or, more cynically, we are going to shoehorn the eurypods into the following theoretical framework):
1. Geographical dispersal: A fairly generalized form radiates geographically without much anatomical change. The innovations introduced by this form, or perhaps some pre-existing mass extinction, creates an environment in which there is little competition. The main constraints are a) environmental limitations and (b) some rather basic anatomical choices. Thus the dinosaurs radiated without much variation from the basic form, except the marginal exception of certain prosauropods, for much of the Middle and Late Triassic. Their anatomy restricted them from marine environments and probably dictated a basic election between carnivorous and herbivorous habits. However, there were few constraints other than such fundamental anatomical and environmental conditions. Note that, so long as the group is expanding its range, selection will tend to disfavor specialization and favor generalist pioneers.
2. Diversification into Guilds: Following consolidation of their position in an ecosystem, the principle constraints become internal to the group. The advantage shifts from highly generalized pioneers to guild specialists who can take advantage of one particular type of environmental resource or lifestyle more efficiently than other members of the group. Like Darwin's finches, this does not require gross changes in body form. Rather the same basic body form is retained, with the development of specific guild-related tools. In the case of dinosaurs, this involved, over the course of the Latest Triassic and Early Jurassic: top predators with larger heads and jaws, high browsers with long necks, middle browsers with considerable range of head motion and specialized dentition, low-browsers with armor and elephantine quadrupedalism -- all retaining the basic dinosaurian body plan.
3. Intra-guild Competition: At some point, particularly with organisms much larger than finches, further guild specialization is not possible. Finches can continue to subdivide ecospace according to seed types almost indefinitely. By contrast, a dinosaur runs out of any specialized resource, within a reasonable geographical range, fairly quickly. This limits the degree of anatomically "easy" specialization possible, unless the beast has very high mobility and access to other geographical pockets with the specialized resource. Arguably, the iguanodont - hadrosaur - marginocephalian group, the Cerapoda, was capable of more specialization (and hence more coexisting similar forms) because of greater mobility. The thyreophorans, burdened by armor, greater weight, and simple graviportal posture, lacked this option. This ecological trap created the conditions for the intense adaptive pressures (and perhaps geographical fragmentation) necessary for major re-engineering of the body plan. That is, further subspecialization could be achieved only by major anatomical changes because of greater size and limited mobility. Hence the basic thyreophoran body types diverged earlier than the corresponding cerapod types. The same could be said of the sauropodomorphs by contrast to the theropods, and for much the same reasons.
Clearly, if one of the new body plans is sufficiently flexible, and conditions are right, this may start a whole new radiation. The radiation of birds from the theropod stem may be an example of this sort of thing. However, for the most part, only a few, increasingly aberrant and specialized forms will survive -- until the specialization becomes so extreme that a change in environment extinguishes the entire line. ATW 011216
|
| Scutellosaurus dawleri Owen (1868) from the Pliensbachian Early Jurassic) of Arizona. About 120 cm and 10 kg. |
Although the closely related Scelidosaurus is sometimes regarded as a primitive ankylosauromorph, and the family is treated in that fashion here. However, Scutellosaurus dawleri, another traditional scelidosaur, probably is truly basal and pre-dates the split between stegosaurs and ankylosaurs. Scutellosaurus was a small lightly built ancestral armoured type. It was a fast sprinter, relying on agility to escape predators, with a light covering of armour plates (scutes) as well. It was originally classified with the fabrosaurs, from which it is no doubt descended. Echinodon becklessi, discussed above, may be another member of this group. MAK990714 & ATW020419.
 Thyreophora: Definition: (stegosaurs, ankylosaurs) > Cerapoda. [C97].
Thyreophora: Definition: (stegosaurs, ankylosaurs) > Cerapoda. [C97].
Genasauria : (Agilisaurus + Cerapoda) + * : Echinodon + (Scutellosaurus + Eurypoda).
fairly weak coronoid process [R56]; $ postorbital process of jugal expanded transversely; broadened pelvis; $ keeled scutes on dorsal surface in parallel rows.
Notes: Scutellosaurus is the most basal form known from reasonably good remains[C97]. It looks more or less like any other advanced dinosauriform, i.e. somewhat bipedal, long necked and long-tailed, but stouter and with the beginnings of armor. ATW011201. Another extremely primitive form, and not so well-known, is Tatisaurus oehleri Simmons 1965. Tatisaurus is an isolated dentary from the Dark Red Beds, Lower Lufeng Series, Yunnan, China, and is probably of Late Hettangian- or Early Sinemurian age (Earliest Jurassic). Tatisaurus is the earliest known member of the Thyreophora It is still not very far removed from its fabrosaur-like ancestors, but within a few million years Tatisaurus, or a form like it, evolved into large armoured quadrupeds, the scelidosaurs.
MAK990714.
Image: Stegosaurus.
Links: DinoData: Thyreophora; Prehistoric World Images Dilophosaurus & Scutellosaurus; Thyreophora - EnchantedLearning.com; Britannica.com; New Page 5; Introduction to the Thyreophora; GEOL 104 Lecture 18- Thyreophora- Defense! Defense! Defense!; Witmer's Lab Dinosaur Skull Collection- Thyreophora; The Armored Dinosaurs by Kenneth Carpenter (ed.); Chapter 14 Links links).
References: Carpenter 1997a) [C97]; Romer (1956) [R56]. ATW030717
Range: Late Jurassic (Tithonian) of North America.
Phylogeny: Thyreophora:: Eurypoda + *.
A bigger (120cm), heavier version of Echinodon.
Range: Early Jurassic Toarcian) of Europe
Phylogeny: Thyreophora : Eurypoda + *.
This animal has been compared to a miniature version of Huayangosaurus, the skull of which is very similiar. MAK990724.
Eurypoda: defined as Stegosaurus + Ankylosaurus
Range: Middle Jurassic to Late Cretaceous. Cosmopolitan.
Phylogeny: Thyreophora :: Scutellosaurus + * : Stegosauria + Ankylosauromorpha
Characters: skull width very broad with intraorbital width at the supraorbitals widest portion of the skull; $ reduced or absent antorbital fenestra and fossa; $ three palpebrals "supraorbitals") form dorsal rim of orbit [G90]; $ supraorbital horns; $ quadrate with shaft and pterygoid ramus in same plane (??) and no distinct lateral ramus [G90]; $ quadrate with articular surface directed ventrolaterally [G90]; $ no "otic") notch between paroccipital process and quadrate [G90]; $ exoccipital ventral portion around foramen magnum recessed and overhung by dorsal part and supraoccipital [G90]; $ deep median keel on hard palate (may have supported soft secondary palate) from vomers & pterygoids [G90]; $ ossified epipterygoid; $ atlas neural arches fused to atlas intercentrum [G90]; parascapular spines present; $ scapular blade of roughly constant width [G90]; robust ulna with convex external margin and huge olecranal expanse; $ short metacarpals with hoof-like unguals (also on pes) [G90]; $ laterally tilted, dorsoventrally compressed preacetabular blade (of ilium, presumably); $ ilium anterior process diverges at least 35° laterally from midline [G90]; ilia broad and overhang entire pelvic area; $ ilium posterior process very short, with antitrochanter (sensu Romer) [G90]; $ partially closed acetabulum (plesiomorphic?); femur neckless, with head almost terminal [G90]; $ enlarged greater trochanter confluent with femoral head; femur with reduction of the fourth trochanter; $ pendent fourth trochanter (?!); mediolaterally expanded tibia with width about equal to craniocaudal length of proximal end; $ proximal tarsals tend to fuse with epipodials in adults [G90]; mediolaterally expanded calcaneum; metapodials short & robust; $ metatarsals short and arranged for spreading, elephantine stance (vs. compact) [G90] [S99]; $ cervical dermal rings (not sure what this means); $ cranial dermal ossicles; parasagittal arrangement of armor.
Links: DinoData: Eurypoda.
References: Galton (1990) [G90]; Sereno (1999) [S99]. 011130.
Ankylosauromorpha:Ankylosaurus > Stegosaurus? But see Ankylosauromorpha Scelidosaurus + Ankylosauria).
Range: Early Jurassic to Late Cretaceous
Phylogeny: Eurypoda: Stegosauria + * : Scelidosauridae + (Minmi + Ankylosauria).
Characters: skull surface extensively modified by reworking bone surface by overlying scales, or by fusion of armor; body very wide (probable hindgut fermentation); 7-8 cervicals, ~16 dorsals, 3-4 sacrals, 40+ caudals; neck short; trunk very long; caudals typically short near hip & elongated near middle of tail; quadrupedal; ilium expanded horizontally & angled away from midline (allows wide hip to accommodate gut); pubes rotated to at least partially close acetabulum; armor over whole body; armor embedded in skin; armor of projecting spines, large narrower spikes, thin walled cone-like plates, keeled nearly flat plates, solid, keeled scutes, with small rounded ossicles on ventral surface & filling gaps; armor in species-specific pattern.
Links: Ankylosauromorpha.
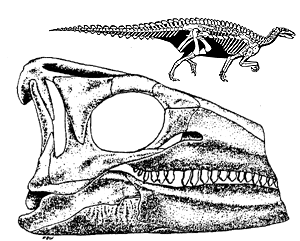 |
| Scelidosaurus skull and skeletal reconstruction from Coombs et al.. (1990), figs. 20.1a & 20.2a. Skeletal reconstruction by Greg Paul. |
Scelidosauridae: Bienosaurus, Scelidosaurus. Early, ankylosaur(-like?) Thyreophorans
Range: Early Jurassic, possibly surviving to Late Jurassic of England.
Phylogeny: Ankylosauromorpha : (Minmi + Ankylosauria) + *.
Characters: Characters: Skull tall posteriorly, but tapering anteriorly to form wedge-shaped lateral profile [C+90]; skull also tapers slightly laterally [C+90]; skull retains all 5 pairs of fenestrae, unlike ankylosaurs; premaxilla with teeth [C+90]; maxilla triangular [C+90]; maxilla with shelf lateral to tooth row [C+90]; nasal long [C+90]; lacrimal rectangular, forming most of anterior orbit [C+90]; nasal-maxillary suture continuous with prefrontal-lacrimal suture [C+90]; additional osteoderm incorporated into orbital rim; jugal and postorbital with extensive overlap, forming broad pillar between orbit and lower temporal fenestra [C+90]; lower temporal fenestra very narrow [C+90]; rugose areas for osteoderms on skull roof, jaws; parietals hour-glass shaped, with narrow waist between upper temporal fenestrae [C+90]; squamosal forms posterior & lateral borders of circular upper temporal fenestrae [C+90]; quadrate vertical, with jaw articulation ventral to tooth row [C+90]; mandibular symphysis broad, rugose & inclined anteriorly (probably a small beak) [C+90]; "cropping beak" of ankylosaurs absent (selective feeder?); predentary unknown or absent [C+90]; mandibular symphysis absent [C+90]; surangular large and forms posterior half of coronoid process [C+90]; spliner-like splenial on ventral margin of jaw [C+90]; teeth with symmetrical enamel [C+90]; tooth roots long and straight [C+90]; vertebral count 6-7 cervicals, 17 dorsals, 4 sacrals, 35 caudals [C+90]; axis centrum fused to atlas [C+90]; neural spines laterally compressed & short [C+90]; parapophyses shift dorsally down the spine, so that last few ribs have single head articulating distally on the transverse process [C+90]; scapular blade short & wide [C+90]; humerus straight, with large, long deltopectoral crest [C+90]; broadening of pelvis; pubes thin & distal pubes curve ventrally away from ischium [C+90]; ilium long post-acetabular region; reversion to quadrupedalism; femur straight to slightly curved [C+90]; femoral head displaced medially, but without a neck [C+90]; lesser trochanter prominent & separated from greater by groove; 4th trochanter large, pendent and placed at mid-length [C+90]; tibia slightly longer than femur [C+90]; large cnemial crest [C+90]; large, free astragalus, large calcaneum and (at least?) 2 other tarsals [C+90]; 4 massive metatarsals & vestigial V [C+90]; phalanges short & flat with blunt, flat unguals [C+90]; phalangeal formula 23450 [C+90]; three rows of plates on each side of body; four rows on each side of the tail; unique 3-pointed plates just behind skull (shoulder spines?) [C+90].
Links: DinoData Classification Scelidosauridae; STEGOSAURS, ANKYLOSAURS (October 8); Scelidosaurus; Lecture 12 - Early Jurassic; dinocountup01.html see entry for Bienosaurus); Scelidosaurus The Natural History Museum's Dino Directory; _Bienosaurus_ questions.
References: Coombs et al. (1990) [C+90]. ATW030606.
checked ATW060215, last modified MAK120308
