Ornithischia
Taxa on This Page
- Agilisaurus X
- Eocursor X
- Fabrosauridae (Lesothosaurus) X
- Genasauria X
- Ornithischia X
Pisanosaurus
"Fabrosauridae" and Other Basal Ornithischians
The cladistic paradigm has meant these early types have been stuck as "outgroups" with no Linnean status beyond family level. Since these early groups differ at least as much as the later ones I decided to take matters into my own hands and christen a new suborder [ Stanley Friesen's Dinosaur classification follows a similar tack].
Fabrosauria - small early ancestral forms - active fast-running lightly-built bipeds, paraphyletic and ancestral to all other Ornithischian lineages, Late Triassic to Early Jurassic - length about a metre. This isn't shown on our cladogram for the obvious reason that it isn't a clade. It collectively refers to things like Lesothosaurus and Agilisaurus. These actually may form a monophyletic group, the Fabrosauridae. In addition, it would include the South American Pisanosaurus, which is probably not a fabrosaurid. (MAK000117 & ATW020420)
The most basal, generally recognized clade is Genasauria. Genasauria is defined as the last common ancestor of Thyreophora and Cerapoda and all of its descendants. The thyreophorans are the armored ornithischians: the stegosaurs and ankylosaurs. The cerapods include both the hadrosaur-like forms and the line which includes both Triceratops and the bone-headed Pachycephalosauria. In short, Genasauria includes all of the major Ornithischian groups.
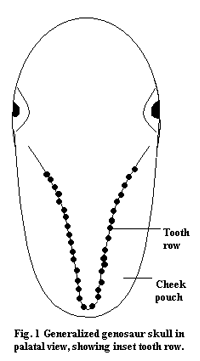 The most important unique characteristic of the group is the inset tooth row. That is, the teeth are not located in the same plane as the sides of the skull. Instead, they are moved in close together, toward the center line. The functional significance of this structure is the advantage it gives to a plant-eater. Herbivores on the model of a sauropod, or even an aetosaur, gulp masses of vegetation. However, leaves, stems and seeds are tough stuff, and chemically hard to digest. This kind of herbivory requires an enormous amount of internal processing, which means carrying around grinding stones in the gut gastroliths) and a big fermentation vat for chemical decomposition. The simple adaptation involved in putting the teeth close together, combined with more teeth and articulating dentition (teeth which are in contact when the mouth is closed and thus cut or crush food between opposing jaws) allows the animal to cut plant food up into tiny bits and crush the juices out of leaves and fruit. This, in turn allows it to do without gastroliths (intestinal rocks for cutting up bulk vegetation), and also to reduce the size of the fermentation area for chemical digestion. Since the pieces swallowed are small, they have a greater surface area per unit volume and can be attacked more quickly by digestive enzymes. This huge gain in efficiency translates into an ability to be more selective about food, to move faster or to carry armor, to spend more time on mate selection or in rearing young. Quite possibly, this simple anatomical change was ultimately responsible for the worldwide relative decline of the sauropods and the extinction of the prosauropods by the Late Jurassic.
The most important unique characteristic of the group is the inset tooth row. That is, the teeth are not located in the same plane as the sides of the skull. Instead, they are moved in close together, toward the center line. The functional significance of this structure is the advantage it gives to a plant-eater. Herbivores on the model of a sauropod, or even an aetosaur, gulp masses of vegetation. However, leaves, stems and seeds are tough stuff, and chemically hard to digest. This kind of herbivory requires an enormous amount of internal processing, which means carrying around grinding stones in the gut gastroliths) and a big fermentation vat for chemical decomposition. The simple adaptation involved in putting the teeth close together, combined with more teeth and articulating dentition (teeth which are in contact when the mouth is closed and thus cut or crush food between opposing jaws) allows the animal to cut plant food up into tiny bits and crush the juices out of leaves and fruit. This, in turn allows it to do without gastroliths (intestinal rocks for cutting up bulk vegetation), and also to reduce the size of the fermentation area for chemical digestion. Since the pieces swallowed are small, they have a greater surface area per unit volume and can be attacked more quickly by digestive enzymes. This huge gain in efficiency translates into an ability to be more selective about food, to move faster or to carry armor, to spend more time on mate selection or in rearing young. Quite possibly, this simple anatomical change was ultimately responsible for the worldwide relative decline of the sauropods and the extinction of the prosauropods by the Late Jurassic.
Of course, narrow jaws are not enough. Other changes were important. For example, if the teeth were merely moved inward, plant material might simply fall out the sides as the dinosaur chewed -- hugely wasteful as well as frustrating. The narrowing of the jaw gives an advantage to an animal with cheeks to keep the food in. Thus, the evolution of genasaurs required cheeks as well as jaw structure innovations. But cheeks, in turn, allow further changes: improvements in jaw musculature, the ability to use salivary enzymes to start chemical digestion in the mouth (not to mention the improved etiquette of chewing with the mouth closed!), reduced water loss by evaporation from the mouth, and so on.
In this connection, it might be useful to examine the details of a basal genasaur. It might -- if there were any to examine. As it turns out, there are probably none. The only form that comes reasonably close is Echinodon. Unfortunately, Echinodon is a Late Jurassic (or even later) species, and most of the relevant developments are presumed to have taken place in the Triassic. Worse, there are few remains. The best Echinodon material is still a handful of disarticulated jaw parts from England originally described by Owen in 1861 and more recently by Galton (1978). Other than this, there is almost nothing. A single tooth from Spain has been referred to the genus. Estes & deAvalle (1982). Callison & Quimby (1984) have discussed post-cranial material from an unnamed and undescribed "fabrosaur" from the Jurassic of Colorado. Olshevsky (1994) and others have suggested that this, and accompanying jaw material, is Echinodon. However, without any attempt by anyone at formal publication of the fossils, it seems pointless to speculate.
As if to emphasize the ambiguity of these bits and pieces, there has been little agreement on the position of Echinodon. Galton (1978) assigns it to the Fabrosauridae. Sereno (1991) disputes a number of important details of Galton's re-description, disputes the very existence of the Fabrosauridae, and suggests that Echinodon is a heterodontosaur. Olshevsky (1994) agrees (more or less), but his conclusions about the phylogenetic place of heterodontosaurs are markedly different from those of Sereno. Coombs et al.. (1990) have an entirely different understanding, and place Echinodon near Scutellosaurus in the Thyreophora, based partly on the association of the English fossils with dermal scutes of uncertain provenance. Perhaps Sereno would now agree. Sereno 1999) currently places virtually all of the fabrosaur material in the basal Thyreophora, although he omits any discussion of Echinodon itself in the most recent study. We, also, treat Echinodon as a thyreophoran; but the choice is rather arbitrary. Much of the debate centers on the "canine" heterodont teeth of the English fossils; but, based on the photographs in Galton's paper, it is not at all certain that the fossil even has such teeth. There are negative casts which might reflect caniniform teeth. In addition, one specimen bears a disarticulated caniniform tooth which might have belonged to Echinodon, but could also have been left by a scavenger. The Fruita specimen is said to have unambiguous fangs, but the specimen remains undescribed.
This is obviously an unsatisfactory state of affairs. In the last analysis, it appears that Echinodon is simply another one of those fossils "whose charm exceeds their scientific merit." Feduccia 1999), quoting Storrs Olsen. It is particularly discouraging that acute and experienced observers such as Sereno and Galton cannot even agree on a description of the known material, much less on its interpretation. All things considered, this is a genus best tucked back into the deep recesses of the British Museum until something more informative comes along. ATW000514.
Descriptions
Ornithischia:Pisanosaurus. All Dinosaurs closer to Triceratops than to birds.
Characters: Cheek teeth with low, subtriangular crowns; teeth foliate with constricted roots; predentary; toothless & rugose snout (probable horn beak); jaw joint below upper tooth row; palpebral; 3 to 5 sacral vertebrae; retroverted pubis; ossified tendons above sacrum; pes V reduced to splint. All had cheeks, horny beaks and were herbivorous. Great variety of "defensive" ornamentation.
Links: ; link
Pisanosaurus mertii Casamiquela, 1967
Horizon: Ischigualasto Formation (Late Carnian) of Argentina.
Phylogeny: basal Ornithischia
Comments: Based on a single, fragmentary skeleton, this is the oldest known, and also the most primitive, ornithischian. The sacrum and orientation of the pubis are not certain, but seem to be propubic, like classic saurischians (Thescelosaurus). The animal when alive was about a meter long MAK120309
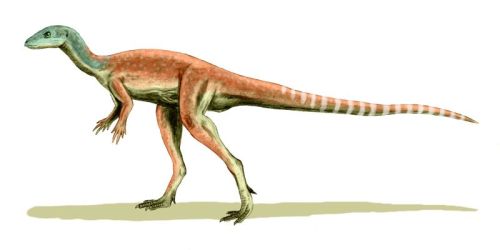
Eocursor parvus Butler, Smith, and Norman, 2007
Horizon: Lower Elliot Formation (Norian) of Orange Free State, South Africa.
Phylogeny: Ornithischia : Pisanosaurus + ((Fabrosauridae /Lesothosaurus + Genasauria) + * )
Comments: Apart from Pisanosaurus this is the oldest known ornithischian. Known from a single specimen comprising substantial cranial and postcranial material, it is also the only Triassic ornithischian known from reasonably complete remains. This was a small lightly built long-legged bipedal dinosaur about 1 meter in length, similar in appaerance to early Jurassic ornithischians such as Lesothosaurus and Scutellosaurus. The tibia is significantly longer than the femur, indicating highly cursorial (fast running) ability. The hands are large and resemble those of the Heterodontosauridae. Providing the earliest evidence for the acquisition of many key ornithischian postcranial characters, including an opisthopubic pelvis. Triangular iguana-like teeth suggest a herbivorous or partially herbivorous diet. Extraordinarily, phylogenetic analysis places Eocursor above the heterodontosaurs, which are here placed near the base of Ornithischia, but below fabrosaurs, and forming a sister taxon to the the Genasauria. The implication is that a large grasping hand is a primitive (plesiomorphic) ornithischian characteristic (Butler et al. 2007). On the other hand, analysis of the basal ceratopsian Yinlong shows the heterodontosaurs to be sister clade to the Marginocephalia (Xu et al. 2006). Assuming the latter (and large manus and other features hared with heterodontosaurs as convergences due to a similar foraging lifestyle), Pisanosaurus, Eocursor, Lesothosaurus represent the fundamental ornithischian condition, from which heterodontosaurs, scelidosaurs, and hypsilophodonts evolved and radiated out from during the early to mid Jurassic. MAK120308
Image: Life reconstruction of Eocursor parvus. Artwork by Nobu Tamura, via Wikipedia, GNU Free Documentation/Creative Commons Attribution Share Alike
 Fabrosauridae: Lesothosaurus (= Fabrosaurus), Agilisaurus, and assorted taxa
Fabrosauridae: Lesothosaurus (= Fabrosaurus), Agilisaurus, and assorted taxa
Range: Late Triassic to Late Jurassic, possibly later.
Phylogeny: Ornithischia : Pisanosaurus + (Eocursor + (Genasauria + *)) (for Lesothosaurus)
Characters: Generally 1-2 m; very lightly built & clearly bipedal; triangular skull; small antorbital fenestra partially occluded by maxilla (primitive); very large orbits & palpebral; lacrimal inserts into slot in maxilla; premaxillary teeth small, laterally compressed, leaf-shaped incisiforms; dentary and maxillary dentition marginal primitive); single wear facets on teeth (vertical tooth action) (primitive); upper and lower teeth occlude alternately; teeth with enamel on medial & lateral surfaces; supra-acetabular buttress over anterior half of acetabulum; femur with head not strongly inturned; femoral shaft slightly bowed anteriorly; fourth trochanter long (caudofemoralis attachment); tibia longer than femur; fibula is reduced to narrow splint; astralagus & calcaneum strongly integrated into tibia and fibula function as part of crus; distal tarsals separate but closely associated with heads of metatarsals; pes tridactyl, with fifth digit lost & first reduced with a slight turn to the rear.
Notes: This likely not a clade, but a paraphyletic collection of very basal ornithischians. The description above largely relates to Fabrosaurus and should not be taken too seriously. ATW
Comment: Fabrosaurus and Stormbergia would seem to be synonyms of Lesothosaurus. A number of poorly-known forms have bene assigned to the Fabrosauridae, including Alocodon, Gongbusaurus, Lufengocephalus, Nanosaurus, Technosaurus, and Xiaosaurus. have been assigned to this group. Some of these, such as Technosaurus, aren't even dinosaurs; others could belong anywhere at around the base or even further up the ornithischian family tree. Middle or Late Jurassic Chinese forms such as Agilisaurus and Gongubusaurus are "fabrosaurs" that represent an advance over Lesothosaurus MAK120308
Links: Literature - Basal Ornithischians & Fabrosaurids; Lesothosaurus - Enchanted Learning Software; Ornithischian dinosaurs: Fabrosaurids; Camptosaurus Nanum (??); Camptosaurus Nanum (view 2); Xiaosaurus - Enchanted Learning Software; FPDM : Agilisaurus louderbacki. ATW020901.
Genasauria: Definition: Thyreophora + Cerapoda.
Range: Early Jurassic to Late Cretaceous
Tooth rows offset medially, from which existence of muscular cheeks is inferred; mandibular foramen reduced and spout-shaped.
Links: DinoData: Genasauria; Lectures 19-20: Late Cretaceous (cladogram); NEW STUDIES; columbia 12; ornithischia; Lecture 19- Late Cretaceous I; ornithischia cladogram; Dinosauria Translation and Pronunciation Guide G. ATW021129.

Agilisaurus louderbacki Peng, 1990
Horizon: Lower Shaximiao Formation of Sichuan, China (Bathonian/Callovian)
Comments: A small, lightly built, cursorial ornithischian of a typically fabrosaur mould. A single complete skeleton is known, making it one of the most complete small ornithischians. agility suggested by its lightweight skeleton and long legs. Its tibia (lower leg bone) was longer than its femur (upper leg bone), which indicates that it was an extremely fast bipedal runner (Wikipedia). The contemporary Hexinlusaurus is similar, although, with a longer tibia relative to its femur, would have been even more agile Thescelosaurus Comparisons between the scleral rings of Agilisaurus and modern birds and reptiles suggest that it may have been diurnal, unlike larger herbivorous dinosaurs that were inferred to be cathemeral, active throughout the day at short intervals (Schmitz & Motani 2011, via wikipedia). Despite the completeness of its remains, Agilisaurus' phylogenetic position is uncertain. Cladistic analyses by Weishampel et al.. 2003; Norman et al.. 2004) found it to be the most basal member of the group Euornithopoda (all ornithopods more derived than Heterodontosauridae). More recent analysis by Zheng et al. 2009 Butler et al. 2011 place Agilisaurus and Hexinlusaurus as stem Cerapoda, an interpretation we have followed here until something better comes along. Regardless of where they belong on the ornithischian family tree, These small and active animals clearly represented an important element of the small herbivore niche, before being supplanted by hypsilophodonts in the late Jurassic. MAK120308
Image Agilisaurus louderbacki, skeleton reconstruction, photo © from Dinosaur Catalogue and Fossil and Mineral World
checked ATW060215, revised MAK120308


