Ecdysozoa
Taxa on This Page
- Ecdysozoa

The diversity of Ecdysozoa, by The Dinosaur Fan, showing a random sleection from each of the eight major phyla (clockwise from lower left): Onychophora, Priapulida, Kinorhyncha, Nematomorpha, Loricifera, Nematoda, Tardigrada, and Arthropoda. Although today these phyla are very distinct, during the Cambrian period there were many intermedate and transitional forms linking them. |
The Evolution of the Ecdysozoa
The Ecdysozoa or "moulting animals" include a number of phyla: Arthropoda, Onychophora, Tardigrada, Kinorhyncha, Priapulida, Loricifera, Nematoda and Nematomorpha that today are very distinct (see collage. above) and have in the past been classified among very different groups of animals. Whilst it is evident on both molecular and morphological grounds that they are related and form a single clade, exact relations between the various phyla remain unclear, reflecting the very rapid evolutionary radiation that occured with the Cambrian explosion.
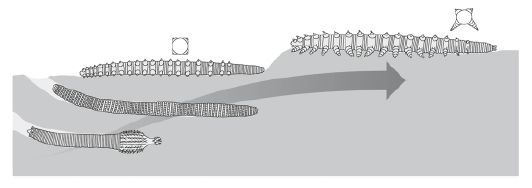
The original Ecdysozoan was probably a largish worm-like organism with a centrally located mouth (Budd 2001). Presumably these would have been infaunal burrowing forms, but some later would have developed legs for greater mobility on the surface. The diagram at the right presents one possible ecological and functional scenario for the evolutionary emergence of ancestral panarthropods (lobopods) from worm-like palaeoscolecid or priapulid-like ancestors (. Other less specialised types would have remained as burrowing worms, while some took up a parasitic lifestyle and became the miniaturised nematoid group
And although the current (extant) ecdysozoan phyla are each very unique and different, this was not the case during the early Paleozoic, especially during the Cambrian, when there were so many transitional and intermediate forms - worms with protoarthropod armour, Dinocaridida that were neither onychophores nor arthropods but somewhere in between, and various forms such as Markuelia and Mureropodia that don't fit any extant phylum. To a Cambrian observer, the Ecdysozoa would have seemed like a single large and diverse, yet still coherent and clearly interrelated phylum, perhaps very much like the Mollusca today.
The ecological composition was also different; for example the palaeoscolecid worms filled many of the ecological roles now taken up by annelids. The anomalocarids filled the apex preditor nich that was since taken up first by large cephalopods and eurypterids, and then by large fish. Some ecolomorphs, such as the soft-bodied and the armoured walking worms, don't seem to have any modern equivalents at all.
At some point, perhaps the Ordovician biodiversification event, this extraordinary range of organisms was pruned, with only a few very distinct lineages surviving. A few holdovers survived to the Silurian and Devonian, but it seems that by the Carboniferous the bulk of taxa belonged to extant groups. Some of these groups, such as the arthropods and the nematodes, became incredibly succssful, others persisted as relics, represented today (and presumably during the Cenozoic and Mesozoic) by a few dozen or a few hundred species.
A similar thing seems to have happened with many taxa during this time, such as molluscs and arachnomorph and crustaceomorph arthropods. One of the reason why phylogeny is so difficult today is that we have so few intermediate types to work with; this is why neontological studies alone can be misleading. Even though fossils don't provide as much data as living species, they do help to show the great diversity of life in the past. MAK120421
Changing phylogenies regarding the Ecdysozoa
The phyla that constitute the ecdysozoa have in the past been classified among very different groups of animals. The Kinorhyncha, Priapulida, Nematoda and Nematomorpha lack a true body cavity - they are pseudocoleomates rather than coelomates - and so were included with the rotifers and gastrotrichs. In other words, arthropods were considered more closely related to vertebrates and molluscs (all three being colemates) than to nematodes. The animal kingdom was largely organised according to a few basic parameters, such as radial and bilateral symmetry, and in the case of the latter, no body cavity (flatworms) versus false (psedocolemates / aschelminthes) versus true (colemate) body cavity (e.g. Margulis and Schwartz, 1982 for a recent version thereof). Similarily, the relation of the colemate phyla was determined by the most apparent similarities. So for example arthropods and onychophores were likewise considered together, along with the annelids, as all have a segmented or quasi-segmented body, and hence all were included under the superphylum Articulata (see e.g. the ideas of Snodgrass).
All this changed with the rise of molecular phylogeny. Though our knowledge of the Ecdysozoa is less than two decades old, this has proved to be a rich field of study, with many changing phylogenetic hypotheses that only now seem to converging on a more detailed consenus. A few of the changing interpretations are shown here on this page
The early interpretations
The Ecdysozoa are such an important concept in metazoan phylogeny that it is hard to imagine that they were only first discovered 15 years ago (at the time of writing). They were first described by Aguinaldo et. al. 1997 who divided the protostomes into the two clades, Ecdysozoa and Lophotrochozoa. He presented the following molecular-based phylogeny
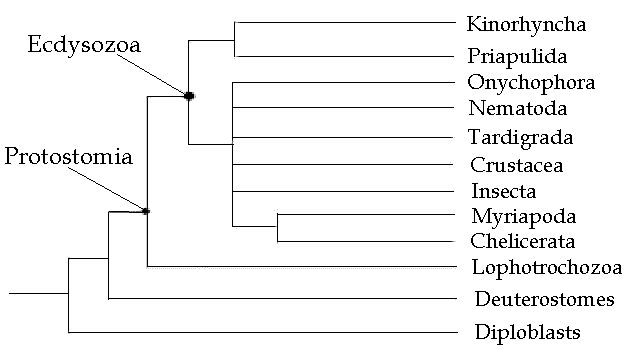
Giribet & Ribera (1998), using 18S rDNA sequences also divide the protostome coelomates into two clades, the Ecdysozoa and the Spiralia. They used DNA sequencing as an alternative to morphology in determining Protostome phylogeny. From their abstract:
"Here we report a cladistic approach to metazoan evolution including 133 18S rDNA sequences of 31 animal phyla. Despite the difficulties associated with analyzing large data sets, our data suggest that the Bilateria and Protostomia are monophyletic. The internal phylogeny of the protostomes is divided into two main clades. One clade includes the classical protostome worms (annelids, sipunculans, echiurans, pogonophorans, and vestimentiferans), mollusks, nemerteans, "lophophorates," platyhelminths, rotiferans, and acanthocephalans, although the internal resolution of the clade is very low. The second clade includes arthropods and other molting animals: tardigrades, onychophorans, nematodes, nematomorphans, kinorhynchs, and priapulans. The arthropods and related phyla lack a ciliated larvae, lack a multiciliate (locomotory) epithelium, and share many features, notably, a reduced coelomic cavity and the presence of a cuticle which molts."
Also this fits in with the modular nature of the Hox cluster of genes, which determine the overall ground plan of an animal. In The Ancestry of the Hox Cluster Guillaume Balavoine writes (from the abstract):
"The 18S rDNA phylogeny of metazoans leads to the picture that the bulk of the bilaterian phyla is split into a double dichotomy giving three great superphyla, Deuterostomia, Ecdysozoa (including nematodes) and Lophotrochozoa (including brachiopods). The four-gene cluster in C. elegans must therefore be the remnant of a larger cluster. Comparison of the Hox [cluster of] genes sequences appears to confirm the position of some phyla in this three-branched tree of the Bilateria, notably brachiopods"
A different molecular phylogenetic study by Winnepennickyx et al. (1998) presents a cladogram that doesn't agree with Aguinaldo et al., although it, too, contains a clade made up of priapulids and arthropods. It also contains clade made by phoronids and brachiopods.
In this context, the Ecdysozoa are more or less equivalent to Conway Morris, 1993earlier but now redundant Arthropodomorpha (dendrogram via Mikko's Phylogeny)
Conway Morris' above morphology-based phylogeny follows the traditional (and in the opinion of the present writer very unlikely) hypothesis of Spriggina as some sort of proto-arthropod. But the rapid Cambrian explosion scenario makes it exceedingly unlikely that Spriggina or its fellows are related (even as stem taxa) to amy extant phyla; indeed, it is not certain that these organisms are even metazoa at all
Peterson & Eernisse 2001) arrive at several different trees through a combination of methods. They indicate that the Lophotrochozoa are paraphyletic and hence not a true clade. Interestingly, all but the morphology only approach showed the sponges (Porifera) to be paraphyletic (i.e. that sponges were the direct ancestors of other forms of animal life) rather than a side-branch
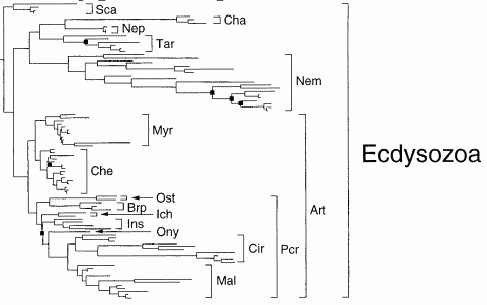
Parsimony analysis of 18s rDNA

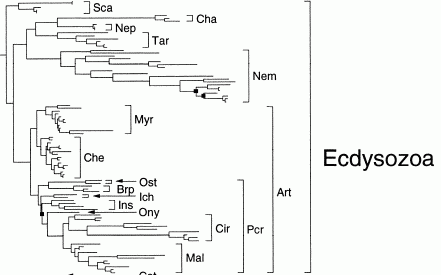
Combined parsimony analysis of morphology and 18s rDNA
The above cladograms by Peterson & Eernisse (2001), show the relationship of the various Ecdysozoa subclades. (Molecular phylogeny). Here tardigrades and chaetognatha are grouped with nematodes
These studies demolished the Articulata, which was shown to be an invalid taxon, with very little morphological, and absolutely no molecular, support. It is clear that both annelids and arthropods acheived a segmented condition independently.
More recent interpretations
As both molecular sequencing and morphology-based cladistics became more refined, two groups emerged as possible subclades of the Ecdysozoa - the Cycloneuralia and the Panarthropoda - and almost as quickly dissappeared into a paraphyletic haze.
The Cycloneuralia are are a large group of sometimes microscopic or miniature mostly wormlike forms, such as the nematodes and kinorhyncha, or larger forms like priapulida. They are distinguished by a collar-shaped brain around the pharynx (also found by convergence in the Gastrotricha, which are lophotrochozoans) and eversible anterior end, or introvert, which terminates in the mouth and gives the alternative name of Introverta Telford et al 2008 p.1531). While often recovered as a monophyletic clade, they have also equally appeared as paraphyletic or as an unresolved polytomy (Telford et al 2008, Edgecombe et al 2011.
The Panarthropoda are a more tightly knit assemblage consisting of the three phyla: Arthropoda, Onychophora and Tardigrada. They share a segmented body plan, the presence of legs and claws, and a ventral nervous system. While the Onychophora and the Arthropoda are clearly related, as shown by both molecular and morphological evidence, the tardigrades' placement is less clear, as in both molecular and morphological details they share characteristics of both panarthropods and cycloneuralia, and can and have been included in either. Indeed, few organisms are more contreoversal than these enigmatic little animals, which have variously been linked with nematodes (Peterson & Eernisse 2001), onychophora (Cavalier-Smith 1998), and arthropods (Budd 2001).
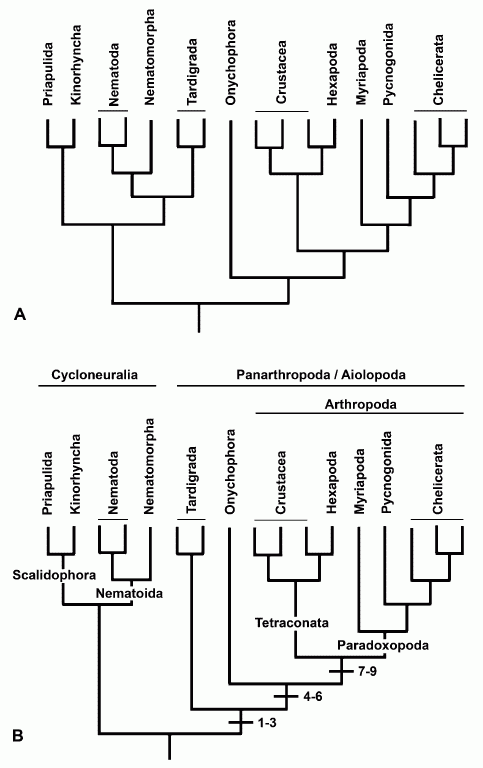
The cladograms on the left present these two alternative hypotheses for ecdysozoan relationships based on molecular data from expressed sequence tags, after Dunn et al. 2008 (see also larger cladogram incorporating phylogeny B) showing the unstable position of the tardigrades. In one analysis (a), tardigrada are placed within a monophyletic Cycloneuralia, in another (b) in a monophyletic panarthropoda, as sister to Onychophora + Arthropoda. Here Cycloneuralia and Panarthropoda are the two sub-clades within the Ecdysozoa. It iis also in accord with cladistic morphological analysis (synapomorphies indicated) whereas the first involves a great deal of parallelism (diagram from Edgecombe 2009 fig.2)
Telford et al 2008, reviews previous work and combines molecular and morphological information to give the following consensual phylogeny
Here the Mandibulata (including Myriapoda) and Panarthropoda (including Tardigrada) are shown as monophyletic groups because of convincing morphological evidence despite some molecular uncertainty.. The Cycloneuralia are now paraphyletic, although the Panarthropoda retain their monophyletic status, even if the relations of the three component phyla remain unresolved.
Rota-Stabelli et al 2010, in addressing arthropod phylogeny from a molecular perspective, sought to avoid conflicting phylogenetic results from molecular datasets by analysing two independent molecular datasets: a phylogenomic dataset of 198 protein-coding genes and new microRNA complements sampled from all major arthropod lineages. The arthropods are considered in the next main unit, but of relevance to the present review, their phylogenomic analysis revealed both a paraphyletic Cycloneuralia and a monophyletic Panarthropoda, suggesting that the previous grouping of tardigrades and nematodes may have been the result of tree-reconstruction artefacts. Interestingly, the Scalidophora (Kinorhyncha + Priapulida) here appear as the sister taxon to an unnamed Nematoda (or presumably Nematoida) + Panarthropoda clade. Tardigrades and onychophores form a monophyletic lobopodia. Campbell et al. 2011 using two independent genomic datasets, arrived at a very similar phylogeny, except that onychophores and arthropods are now sister taxa and showing the tardigrade-nematode group to be a phylogenetic artifact resulting from long branch attraction, due to tardigrades' high rate of evolution. Their analysis strongly supported a monophyletic Panarthropoda including Tardigrada and suggest a sister group relationship between Arthropoda and Onychophora, in keeping with current morphological and fossil evidence. It also revealed a paraphyletic Cycloneuralia and a monophyletic Scalidophora. Their two molecular phylogeny is as follows:
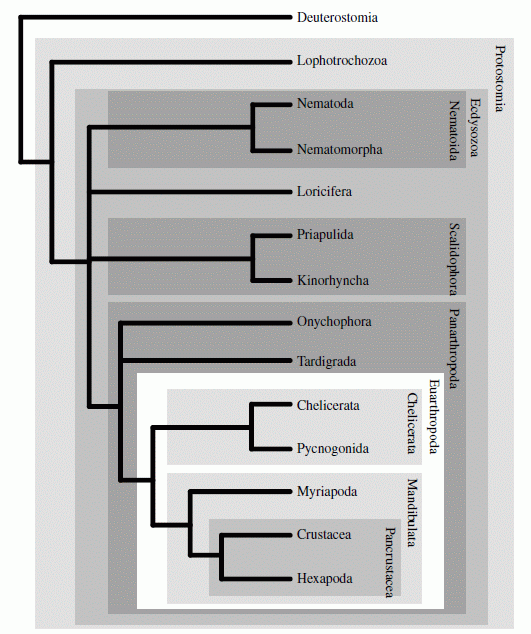
 Phylogenetic tree by < a href="references.html#Campbelletal2011">Campbell et al. 2011, using Bayesian analysis of molecular data, supports a monophyletic panarthropoda with tardigradesas the sister group of Onychophora + Arthropoda (defined as Lobopodia in this paper). This is in agreement with morphological and fossil evidence. Other findings shown here, such as a paraphyletic Cycloneuralia and monophyletic Scalidophora, Mandibulata, and Pancrustacea, are part of an emerging molecular consensus, although the Scalidophora may only be monophyletic as far as the crown group goes. The problem of a molecular and morphological Mandibulata and molecular-only Pancrustacea verses a fossil and morphological Schizoramia and Mandibulata will be addressed under Arthropods (next unit).
Phylogenetic tree by < a href="references.html#Campbelletal2011">Campbell et al. 2011, using Bayesian analysis of molecular data, supports a monophyletic panarthropoda with tardigradesas the sister group of Onychophora + Arthropoda (defined as Lobopodia in this paper). This is in agreement with morphological and fossil evidence. Other findings shown here, such as a paraphyletic Cycloneuralia and monophyletic Scalidophora, Mandibulata, and Pancrustacea, are part of an emerging molecular consensus, although the Scalidophora may only be monophyletic as far as the crown group goes. The problem of a molecular and morphological Mandibulata and molecular-only Pancrustacea verses a fossil and morphological Schizoramia and Mandibulata will be addressed under Arthropods (next unit). |
In the consensus tree suggested by Edgecombe et al 2011, not only are the Cycloneuralia paraphyletic, but the tardigrades are once again unresolved, resulting in the Panarthropoda also becoming a paraphyletic grouping leaving only Onychophora and the Arthropoda in a reduced (and unlabelled) Eu-Panarthropoda. We have however followed the phylogeny of both Molecular phylogeny according to Rota-Stabelli et al 2010 and Campbell et al. 2011, according to which the grouping of tardigrades and nematodes is an artifact of earlier phylogenomic analyses. As molecular phylogeny increasingly addresses the problem of Long Brabnch Attraction, a more accurate phylogenetic tree will certainlty emerge.
Which came first, the worm or the lobopod?
This is a bit like the story of Archaeopteryx and the deinonychosaurs, or perhaps lampreys and ostracoderms. Did evolution follow a simple ascending path of greater complexity and specialisation? Or did the more advanced form evolve first, from earlier unknown primitive ancestors, and then , througha sort of reverse evolutiobn, give rise to more primitive or degenerate (or in the case of Archaeopteryx, flightless longer tailed) descendants?
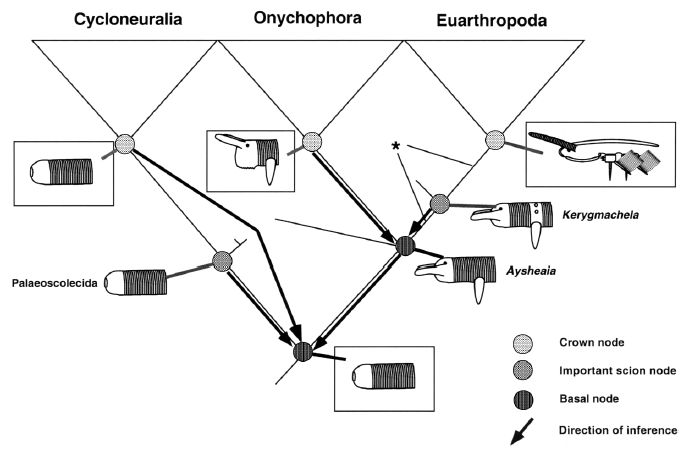 Reconstructing the basal and crown nodes in the Ecdysozoa. Nodes shaded according to key. Important plesions that are essentially metataxa (i.e. their character states cannot presently be distinguished from their respective plesion node) are shown. Reconstructed character states are shown inside boxes. Inference of character states at different nodes shown by bold arrows. Diagram and caption by Budd 2001 Reconstructing the basal and crown nodes in the Ecdysozoa. Nodes shaded according to key. Important plesions that are essentially metataxa (i.e. their character states cannot presently be distinguished from their respective plesion node) are shown. Reconstructed character states are shown inside boxes. Inference of character states at different nodes shown by bold arrows. Diagram and caption by Budd 2001 |
In considering the evolution of the Ecdysozoa, we naturally think that the ancestral form was a worm-like organism. This is illustrated by the cladistic diagram presented by Budd 2001 (left). Here an ancestral, worm-like form (shown as the basal node, at the bottom of the diagram) gives rise to the Cycloneuralia (shown on the left of the diagram) and the panarthropod clades (the Onychophora and Arthropoda are shown on the right; the position of the tardigrades is shown by the asterisk but not more precisely indicated because of its uncertain phylogenetic placement,. A later paper by Budd & Telford (2009), in keeping with developments (and new ambiguities) since the first paper was written, has a low resolution consensus phylogeny, with an unresolved panarthropoda(paraphyletic lobopoda) arising from an unresolved ecdysozoa (paraphyletic cycloneuralia). Our understanding is a bit better now because as we have seen, molecular phylogeny now places onychophores and arthropods as sister taxa, with tardigrades as an early off-shoot. The "worms came first" hypothesis is also This is supported by paleontological research into early armoured worms called, palaeoscolecids as well as lobopods (Dzik & Krumbiegel 1989, Liu et al 2008, Conway Morris & Peel 2010, Steiner et al 2012)
Going in the opposite direction, to more rather than less speculation is a rather exotic phylogeny by Zhuravlev et al 2011, according to which nematoida and related taxa are minaturised and simplied relatives or descendents of a lobopod-like ancestor. The authors provide this richly detailed diagram:
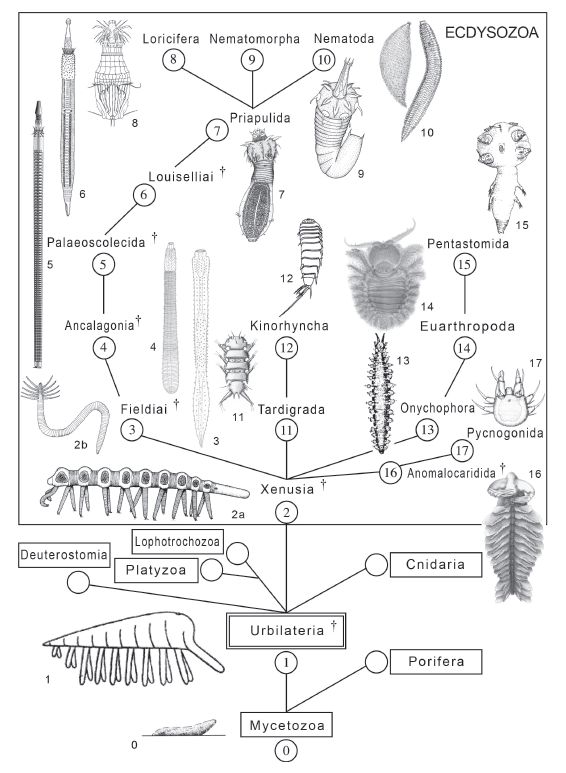 General phylogeny of the Ecdysozoa 0.migration slag of amoebozoan Dictyostelium discoideum Raper, extant; 1.Urbilaterian, hypothetic (modified from van Beneden, 1891); 2a.xenusian Microdictyon sinicum Chen, Hou and Lu, Lower Cambrian (modified from Hou and Bergstrom, 1995); 2b.xenusian Facivermis yunnanicus Hou and Chen, Lower Cambrian (modified from Delle Cave and Simonetta, 1991); 3.Fieldia lanceolata Walcott, Middle Cambrian (modified from Conway Morris, 1977); 4.Ancalagon minor Walcott, Middle Cambrian (modified from Conway Morris, 1977); 5.palaeoscolecid Cricocosmia jinningensis Hou and Sun, Lower Cambrian (modified from Han et al., 2007); 6.Louisella pedunculata Walcott, Middle Cambrian (modified from Conway Morris, 1977); 7.priapulid larva Halicryptus spinulosus von Seibold, extant (modified from Malakhov and Adrianov, 1995); 8.loriciferan larva Pliciloricus ornatus Higgins and Kristensen, extant (modified from Malakhov and Adrianov, 1995); 9.nematomorph larva Gordionus senkovi Malakhov and Spiridonov, extant (modified from Malakhov and Adrianov, 1995); 10.nematodes Greeffiella (left) and Criconema (right), extant (modified from Brusca and Brusca, 2003); 11.tardigrade Stygarctus abornatus McKirdy, Schmidt and McGinty-Bayly, extant (modified from McKirdy et al., 1976); 12.kinorhynch Centroderes eisigii Zelinka, extant (modified from Malakhov and Adrianov, 1995); 13.onychophoran Peripatopsis moseleyi Wood-Mason), extant (modified from Ruhberg in Monge-Najera and Hou, 1999); 14.larval euarthropod Ascalaphus sp., extant; 15.larval pentastomid Boeckelericambria pelturae Walossek and Muller, Upper Cambrian (modified from Maas and Waloszek, 2001); 16.anomalocaridid Anomalocaris saron Hou, Bergstrom and Ahlberg, Lower Cambrian (modified from Hou et al., 1995); 17.protonymphon larva of pycnogonid Anoplodactylus sp., extant (modified from Maxmen et al., 2005).. Caption and diagram by Zhuravlev et al 2011. General phylogeny of the Ecdysozoa 0.migration slag of amoebozoan Dictyostelium discoideum Raper, extant; 1.Urbilaterian, hypothetic (modified from van Beneden, 1891); 2a.xenusian Microdictyon sinicum Chen, Hou and Lu, Lower Cambrian (modified from Hou and Bergstrom, 1995); 2b.xenusian Facivermis yunnanicus Hou and Chen, Lower Cambrian (modified from Delle Cave and Simonetta, 1991); 3.Fieldia lanceolata Walcott, Middle Cambrian (modified from Conway Morris, 1977); 4.Ancalagon minor Walcott, Middle Cambrian (modified from Conway Morris, 1977); 5.palaeoscolecid Cricocosmia jinningensis Hou and Sun, Lower Cambrian (modified from Han et al., 2007); 6.Louisella pedunculata Walcott, Middle Cambrian (modified from Conway Morris, 1977); 7.priapulid larva Halicryptus spinulosus von Seibold, extant (modified from Malakhov and Adrianov, 1995); 8.loriciferan larva Pliciloricus ornatus Higgins and Kristensen, extant (modified from Malakhov and Adrianov, 1995); 9.nematomorph larva Gordionus senkovi Malakhov and Spiridonov, extant (modified from Malakhov and Adrianov, 1995); 10.nematodes Greeffiella (left) and Criconema (right), extant (modified from Brusca and Brusca, 2003); 11.tardigrade Stygarctus abornatus McKirdy, Schmidt and McGinty-Bayly, extant (modified from McKirdy et al., 1976); 12.kinorhynch Centroderes eisigii Zelinka, extant (modified from Malakhov and Adrianov, 1995); 13.onychophoran Peripatopsis moseleyi Wood-Mason), extant (modified from Ruhberg in Monge-Najera and Hou, 1999); 14.larval euarthropod Ascalaphus sp., extant; 15.larval pentastomid Boeckelericambria pelturae Walossek and Muller, Upper Cambrian (modified from Maas and Waloszek, 2001); 16.anomalocaridid Anomalocaris saron Hou, Bergstrom and Ahlberg, Lower Cambrian (modified from Hou et al., 1995); 17.protonymphon larva of pycnogonid Anoplodactylus sp., extant (modified from Maxmen et al., 2005).. Caption and diagram by Zhuravlev et al 2011. |
Although the idea of a lobopodous ur-bilaterian may be rather implausible, the possibility that wormlike ecdysozoan phyla are highly derived (and in the case of nematodes etc miniaturised and simplified) lobopods is not as incredible as it seems. Especially in the case of primitively parasitical nematoida, this would not be very different to the the case of the highly specialised and parasitic pentastomids, once believed to be a distinct phylum, but now almost unanimously agreed to be derived crustaceans. Whether evolution really did go that way regarding the wormlike and mostly microscopic cycloneuralia cannot be known for sure as yet. Although the majority view has the panarthropods as a specialised line (or convergent lines, if the tardigrades are distinct), derived from worm-like ancestors, it may also be that worm-like ecdysozoa gave rise to early lobopods, some of which in turn evolved into neo-worms, whilst other worms - for example the large priapulids which are better understood as primitive burrowers - never went through a lobopodous stage. This would be equivalent to a compromise maniraptoran hypotheiss, in which early coleurosaurs evolved into archaeopterids, some of which became neo-flightless theropods and others advanced birds. For now however we have followed a more traditional phylogeny. MAK120418
Molecular versus fossil phylogenies
It is no doubt obvious that the molecular phylogenies on this page differ quite markedly from the paleontological morphology and ecomorphic-based scenarios. The reasons for this are both methodological and the result of the differeing data each one incorporates. Molecular phylograms use branching trees with no actual ancestors and without reference to stratigraphic, ecological, or functional factors (the same applies to cladograms, hence molecular and morphology-based phylogenies are often grouped together and even synthesised, despite the molecular phylogenies contradicting obvious morphological evidence). Paleontological morphology-based scenarios in contrast examine the fossil record (especially exceptionally preserved fossils) to understand the groundplans and traits of actual ancestral groups, as well as the evolutionary and morphofunctional trends and processes leading to the development and emergence of new types of organisms. Because molecular evolution is based, obviously, only on extant taxa, it is limited to isolated lineages that are generally very far from their ancestral states (the exception would be currently evolving and diverging taxa that are still undergoing an adaptive radiation, e.g. bovid mammals, a very diverse group whose evolution only began in the Miocene). Long branch attraction, homoplasy, and other artifacts may result in the appearance of artificial clades, such as the Cyclostomata among basal vertebrates, the pleistomollusca (gastropods + bivalves) among molluscs or the tetracoronata/pancrustacea (Crustaceans + Insects) among arthropods, clades that in each case are well represented on molecular grounds but make little or no morphological sense. In all these instances there is the assumption that the phylogenetic signal in molecular sequencing is superior to that in morphology-based cladistics or morphofunctional or paleontologfical-morphological approaches, when no empirical evidence has been offered for this (although to be fair neither has any for the converse premise, that molecules have a less reliable phylogenetic signal)
When considering fossil evidence, even from lagerstätten, we need to be sensitive to the fact that very little data remains, relative to the great wealth of data (morphological, genomic, developmental, etc) available for extant taxa. It is easy for the faint phylogenetic signal to be swamped by the neontological noise. This is why additional emphasis should be given to these precious relics and clues from the past. Hence unique transitional taxa like Kerygmachela, Markuelia, Mureropodia, and Sirilorica. The most likely picture that emerges is of a paraphyletic assemblage, or evolutionary grade, of worm-like forms at the base of the ecdysozoan family tree. The isolated extant clades such as cycloneuralia, scalidophora, nematoida, and panarthropda, that emerge through molecular phylogeny are either of a phylogenetically limited sample of recent only forms, or the highly derived descendents of these early worms, or both. And because transitional types such as Mureropodia and Facivermis bridge the gap between scalidophoran worms and panarthropods, it seems likely that Scalidophora and Cycloneuralia, rather than being monophyletic clades, are paraphyetic assemblages (and hence, cladistically speaking, synonyms of Ecdysozoa). In this scenario, both the large and complex lobopods and arthropods on the one hand, and small and simple nematoids on the other, evolved as specialised, albeit astonishingly successful (specifically,m the arthropods and nematodes) side branches from the original priapozoan stem. MAK120422 revised 120429
Descriptions
Ecdysozoa: bugs > slugs
Phylogeny: Protostomia : Lophotrochozoa + * : Scalidophora (syn. of Ecdysozoa?) + (Nematoida + Panarthropoda)
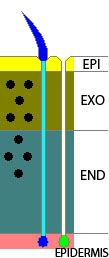
thick three-layered cuticle composed of organic material (see diagram of arthropod cuticle, right), which is periodically molted as the animal grows (ecdysis); no surface locomotory cilia [n1], ciliary feeding mechanisms, or ciliated larvae; generally amoeboid sperm; embryos do not undergo spiral cleavage (Wikipedia); ; gut, if present, with anus ; coelom present only embryonically or absent; ventral nerve cord; usually gonochoristic (Cavalier-Smith (1998)); molting is controlled by characteristic hormones called ecdysteroids; the cuticle has 3 layers: epicuticle, exocuticle, and endocuticle; epicuticle is trilaminate, alpha-chitin in inner layer (Harvey et al. (2010)); primitively, triradiate pharynx (retained in tardigrades, pycnogonids and nematodes), homonomous annulation (priapulids and panarthropoda) (Daley et al 2009 cladogram, supplement fig S3)
Comments: The moulting animals, one of the most successful and diverse clades of life on Earth. First proposed on molecular grounds by Aguinaldo et al 1997; since then a number of morphological synapomorphies have been discovered. Budd tentatively reconstructs the ecdysozoan common ancestor as a large worm-like form with a terminal mouth (Budd 2001); if Cycloneuralia are paraphyletic it may also have possessed a cycloneuralian brain and an introvert feeding mechanism. It is even possibile that the ecdysozoan ancestor was segmented. The similar deployment of homologous genes ("segment polarity" or "pair rule" genes) in arthropods and kinorhynchs would imply common ancestry of segmentation within the group. Telford et al 2008 MAK120415
Note: [n1] Although this would seem to follow from the occurrence of a thick cuticular covering, Telford et al. (2008) and Edgecombe et al. (2011) note that Gastrotricha (a phylum in the Spiralia) does have both a thick cuticle and a body covered by cilia. - Jack R.Holt Diversity of Life - Ecdysozoa
Links Introduction to the Ecdysozoa; Jack R.Holt Diversity of Life - Ecdysozoa; The Ecdysozoa: true taxon or molecular mirage? Brian K. Penney; Ecdysozoa photos; Ecdysozoa - Wikipedia; Reaching creationists: here's the toolbox, do you know how to use the tools? - PZ Myers (includes discussion of Edgecombe 2009)
page uploaded 7 April 2002, last modified 2 May 2003, checked ATW050806, last modified MAK120429. All original content by M. Alan Kazlev Creative Commons Attribution . Other content copyright respective authors or publishers