Crustacea - 1
 This page introduces the crown group Crustacea, sometimes defined as the last common ancestor of all living crustaceans and all of its descendants. This is, of course, a circular definition. We normally try to give a somewhat more explicit definition of a clade, but that's admittedly difficult for the Crustacea. Nothing remotely resembling a consensus phylogeny of the main crustacean groups has emerged from 250 years of carcinology. Martin & Davis 2001). In fact, we even lack agreement on what the "main crustacean groups" might be. For example, a respectable minority holds that Crustacea itself is paraphyletic and includes the insects. See, e.g., Cook et al. 2005); Regier et al. (2005) and discussion below.
This page introduces the crown group Crustacea, sometimes defined as the last common ancestor of all living crustaceans and all of its descendants. This is, of course, a circular definition. We normally try to give a somewhat more explicit definition of a clade, but that's admittedly difficult for the Crustacea. Nothing remotely resembling a consensus phylogeny of the main crustacean groups has emerged from 250 years of carcinology. Martin & Davis 2001). In fact, we even lack agreement on what the "main crustacean groups" might be. For example, a respectable minority holds that Crustacea itself is paraphyletic and includes the insects. See, e.g., Cook et al. 2005); Regier et al. (2005) and discussion below.
The Crustacea are both diverse and common. The usual estimate of 50,000+ known species is probably a gross underestimate. Martin & Davis 2001). Actual diversity is certainly many times that number. The standing crop of just one species, the Antarctic krill Euphausia superba, is said to approach 500 million tons. Id. Indeed, the majority of zooplankton are Crustacea - either larvae or tiny adults. Martin & Davis (2001) quote Geoffrey Fryer for the proposition that the characteristic crustacean nauplius larva is "the most abundant type of multicellular animal on earth."
The vast majority of crustaceans are marine. Crustaceans are typically benthic filter feeders who use various appendages to force water through setae attached to other limbs. Virtually all other modes of life may also be found in any reasonably large taxon of Crustacea. A few (slaters or sow bugs) even live on land, usually under old logs and leaf litter.
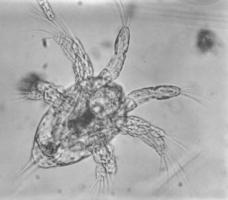 But to really appreciate the Crustacea, you have to see them from a carcinologist's point of view. To the carcinologist, crustaceans are not simply one of the four main arthropod lineages. The carcinologist recognizes that the arthropods hit the Middle Cambrian running and very quickly produced side branches leading to the chelicerates, myriapods, and various extinct groups. Somewhat later, the Hexapoda went off to handle some terrestrial arthropod niches. The Crustacea, however, are the main line and torch-bearer of arthropod evolution; and have carried that torch since the Cambrian "explosion" of metazoans.
But to really appreciate the Crustacea, you have to see them from a carcinologist's point of view. To the carcinologist, crustaceans are not simply one of the four main arthropod lineages. The carcinologist recognizes that the arthropods hit the Middle Cambrian running and very quickly produced side branches leading to the chelicerates, myriapods, and various extinct groups. Somewhat later, the Hexapoda went off to handle some terrestrial arthropod niches. The Crustacea, however, are the main line and torch-bearer of arthropod evolution; and have carried that torch since the Cambrian "explosion" of metazoans.
There is something to be said for this point of view. It isn't the usual parochialism of monomaniacal specialists. Crustaceans have no single body plan, no ubiquitous limb specialization, and no consistent number of segments per major body division. Perhaps that isn't quite true. The crustacean head is fairly stereotyped. However, none of the recent papers we examined would even propose a list of synapomorphies for Crustacea. A good many characteristics are common among Crustaceans, but we are unsure if any are both universal and unique to the Crustacea. If you're interested, it should be possible to derive some synapomorphies from the data in Giribet et al. 2005). In any event, there is a very reasonable case to be made that the arthropods include (a) various small stem groups, mostly extinct, (b) two larger specialized groups, the chelicerates and myriapods, and (c) the insects, with their stem group (Hexapoda). Everything else is Crustacea.
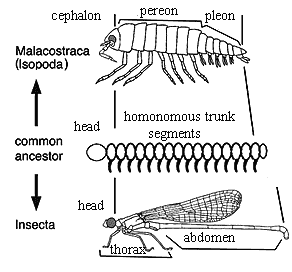 This state of affairs is a recent improvement. Up until a few years ago, it was unclear which of the major arthropod taxa was the sister-group of the Crustacea; but essentially all of the studies published since the late 1990's favor the hexapod-crustacean clade called Pancrustacea or, sometimes, Tetraconata. In fact, as mentioned earlier, it is no longer completely clear that insects are separate from crustaceans. Most (but not quite all) molecular phylogenies in the last few years show Hexapoda as a group within Crustacea. See, for example, Lavrov et al. 2004); Cook et al. (2005); Regier et al. 2005). Then again, some molecular phylogeny can usually be found to support almost any phylogenetic hypothesis, no matter how demented.
This state of affairs is a recent improvement. Up until a few years ago, it was unclear which of the major arthropod taxa was the sister-group of the Crustacea; but essentially all of the studies published since the late 1990's favor the hexapod-crustacean clade called Pancrustacea or, sometimes, Tetraconata. In fact, as mentioned earlier, it is no longer completely clear that insects are separate from crustaceans. Most (but not quite all) molecular phylogenies in the last few years show Hexapoda as a group within Crustacea. See, for example, Lavrov et al. 2004); Cook et al. (2005); Regier et al. 2005). Then again, some molecular phylogeny can usually be found to support almost any phylogenetic hypothesis, no matter how demented.
Both molecules and (to a lesser extent) morphology, also suggest a particular connection between the most successful groups of aquatic Crustacea, the Malacostraca, and the most successful group of terrestrial hexapods, the Insecta. This putative insect/malacostracan relationship is probably a mirage caused by Williston's Law applied to similar patterns of tagmosis. We may get to that issue in more detail below. The important point is that, putting aside these peculiar notions (although we may well come to regret our skepticism), recent reports contain a goodly number synapomorphies which unite the hexapods and crustaceans, to the exclusion of everyone else.
The factor which has propelled this recent flurry of interest (and progress) in Crustacean phylogeny is the maturing business of "evo-devo." After writing that last sentence, we thought we'd better work up a short background piece on the evo-devo of hox genes, since they are important to what follows. That was almost six months ago. Hopefully, you are already a haruspice of hox, or at least have ready access to Carroll (2005). Otherwise, we will be forced to assign you this: remedial hox homework. [2].
| Crustacea: The Usual Suspects |
| Branchiopoda |
Artemia, Daphnia. Arguably the most basal group. |
| Cephalocarida |
|
| Malacostraca |
Crabs, real shrimp, lobster, etc. |
| Maxillopoda |
|
| Ostracoda |
|
| Remipedia |
Blind cave-dwellers, only discovered in the 1970's |
We commonly discuss diversity only after developing some vague notions concerning the sort of organism under discussion. But, Crustacea being a sort of catch-all, we need to offer an example first. If nothing else, this may help you appreciate the size and difficulty of the problem the Crustacea represent. For a more rational taxon, we would start with the most basal of the major included groups. Unfortunately, there is no consensus about who the main groups of crustaceans might be, much less which one is basal. The usual suspects are noted in the table. Any one or more of them may be paraphyletic. Any one or more may be the earliest branching, except possibly Ostracoda. At least, we have never seen a paper in which the claim is made that ostracods are basal to other crustaceans.
Perhaps the most frequently mentioned candidate for basal Crustacea are the Branchiopoda. They have the advantage of possessing a rather good Furongian fossil record. In addition, the branchiopods include 800-900 species of extant fairy shrimp, sea monkeys, tadpole shrimp, water fleas, and clam shrimp. Where do people come up with these asinine names? None of the branchiopods are actually shrimp, of course. Nor do they bear the slightest resemblance to fairies, monkeys, tadpoles, fleas, clams, etc.
One of these extant genera is the "brine shrimp" or "sea monkey," Artemia, which, naturally, is not a shrimp, much less a monkey, and does not live in the sea. Artemia is probably so named because it has no conceivable connection with the goddess Artemis, either. Despite a conspicuous lack of theomorphism, Artemia is scientifically important. Considerable  biological and genetic work has been done with Artemia on the theory that it is a very primitive crustacean. Whether or not Artemia is actually primitive for Crustacea, there seems to be general agreement that the Anostraca in general, and Artemia in particular, are plesiomorphic within Branchiopoda.
biological and genetic work has been done with Artemia on the theory that it is a very primitive crustacean. Whether or not Artemia is actually primitive for Crustacea, there seems to be general agreement that the Anostraca in general, and Artemia in particular, are plesiomorphic within Branchiopoda.
Branchiopods tend to be no larger than 2 cm in length, with the exception of a few "giant" Cladocera, which can reach 10 cm or more. Most branchiopods live in fresh or brackish water. Many, like Artemia, are specialists, adapted to life in temporary pools. They often have the expected suite of adaptations for that lifestyle, e.g. a broad tolerance for changes in salinity (are euryhaline), rapid development, facultative parthenogenesis, and a spore-like resting stage in which the fertilized egg can survive for years, if necessary, until conditions are favorable for development [4].
We will refer frequently to Artemia -- not because it is necessarily either typical or primitive, but because so many people have thought it so. Consequently, it is much studied and relatively well-known. Artemia is not a "typical" crustacean, if there is such a thing. We will use the Branchiopoda, and particularly Artemia, simply as a point of reference, so that we don't waste too much time having to re-orient ourselves to the numerous different geometries which the Crustacea have adopted.
 Arthropods are segmented animals, and Crustacea are no exception. Generally, the segments are organized into three groups, the head (cephalon), thorax (or pereon), and abdomen (or pleon) [1]. These groups are referred to as tagmata. Generally, the Branchiopoda show little tagmosis. The head, thorax, and abdomen are continuous and less markedly differentiated than in most crustaceans.
Arthropods are segmented animals, and Crustacea are no exception. Generally, the segments are organized into three groups, the head (cephalon), thorax (or pereon), and abdomen (or pleon) [1]. These groups are referred to as tagmata. Generally, the Branchiopoda show little tagmosis. The head, thorax, and abdomen are continuous and less markedly differentiated than in most crustaceans.
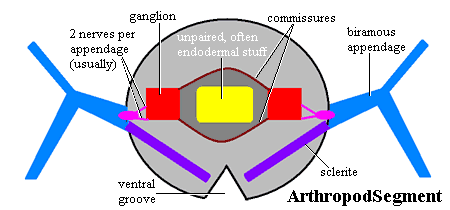
But just what is a segment? For now, we will confine ourselves to the anatomical part of the answer. If one asks about segments, one is usually told about appendages, or about engrailed expression and the process of segmentation, or about hox expression boundaries. What one is rarely told is that the arthropod segment is essentially a neurological unit. But see Harszch 2006). To be more exact, the essence of the arthropod segment seems to be a pair of nerve ganglia attached (a) to each other by one or two commisures, (b) to adjacent segments by a pair of ventral nerve cords (one on each side, or a single fused nerve cord), and (c) to an appendage, if one is present, again usually by two nerves. Arthropods can do almost anything with a segment, but this basic neurological unit is remarkably constant -- from the anterior protocerebrum of primitive pycnogonids (Maxmen et al., 2005) to the telson of derived insects (Richards, 1955). Fanenbruck et al. (2004); Scholtz & Edgecombe (2005).
Leg Words: an Epistemological Endite
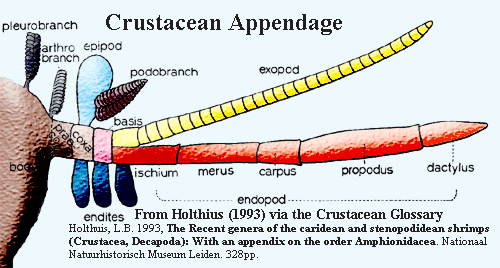
But however comforting the constancy of the arthropod nervous system may be, the appendages get most of the press and require our urgent attention. Almost anything which sticks out of a crustacean, and some things that don't, is a an appendage, and is built on a characteristic appendicular ground plan. For example, the antennae are just specialized legs. In fact, they often begin as larval swimming legs.
Crustaceans are justly famous for the diversity of their appendages, and carcinologists are just as rightly vilified for the complex, inconsistent, terminology they have associated with their legs. We are uncertain how non-carcinologists were expected to deal with the subject before Dr. Joel Martin created the on-line Crustacea Glossary. As a precaution, we have downloaded all 400 pages of Dr. Martin's Glossary -- just in case he should meet with an unfortunate accident after exposing these inner mysteries of carcinology to the profane gaze of the uninitiated.
The fundamental appendage rules are three:
1) The crustacean leg may branch like a particularly aggressive shrub. However, the main axis is conceived to run through two proximal elements called the basis and coxa (basis + coxa = protopod), then through a variable number of distal elements collectively referred to as the endopod see figure).
 (2) The suffixes -ite, -pod, and -podite are all equivalent and are equally meaningless. Thus coxa, coxite, coxapod, and coxapodite all refer to the first segment of an appendage. The suffix -cerite is sometimes used for antennal elements (or articles). The only useful common suffix is -branch, which tells us that the segment is a gill.
(2) The suffixes -ite, -pod, and -podite are all equivalent and are equally meaningless. Thus coxa, coxite, coxapod, and coxapodite all refer to the first segment of an appendage. The suffix -cerite is sometimes used for antennal elements (or articles). The only useful common suffix is -branch, which tells us that the segment is a gill.
3) The prefixes do all the real work. They are best understood as directional indicators. Thus an epipod grows upward, an exopodite points out, and an endite grows down. Other industrious prefixes are positional: arthro- = at the "joint" (i.e., the base) of the leg and pleuro- = off to the side somewhere. Fuzzy Exception: as stated, the endopod is the outer part of the "main" axis. However, it isn't always clear whether an element is an endopod, or simply the outermost member of a series of endites, or whether, in fact, there is no difference and we have once again been duped into making meaningless Latinate noises. See image adapted from Oleson (2007).
For more dark mutterings and skepticism about this entire scheme, see our glossary entry at endopod.
The arthropod head is one of those horrendously difficult problems in homology which we encounter from time to time. On a biological scale of intellectual misery, the cephalon falls somewhere between the fungal cell wall and the vertebrate braincase. This may explain why we have not been able to locate a single, reasonably complete image of the crustacean head. Perhaps a more important reason is that no known crustacean, living or extinct, actually displays a complete set of head parts laid out in such a way that the segmental relationships are clear. In addition, the homologies are contentious. We probably ought to give a fair and balanced review of the topic, but we are not good at that sort of thing. In any case, the fighting is over the chelicerates and stem group arthropods. Budd (2002); Scholtz & Edgecombe (2005). To the extent there is disagreement, we generally follow the system of Scholtz & Edgecombe (2005). Our views have also been corrupted by the work of Prof. Dieter Waloszek [5] in ways which are difficult to tie down with particular citations.
But enough name-dropping. The essential correspondences are listed in the table. We'll work through the head in a generally anteroposterior fashion. This all sounds impressively organized, but no regular reader of Palaeos will be fooled. We will quickly degenerate into the usual random remarks and chaotic speculation. To minimize the inevitable disorientation and distracting nausea, please look carefully at the table before reading the discussion.
Acron: Eyes and Invisible Legs
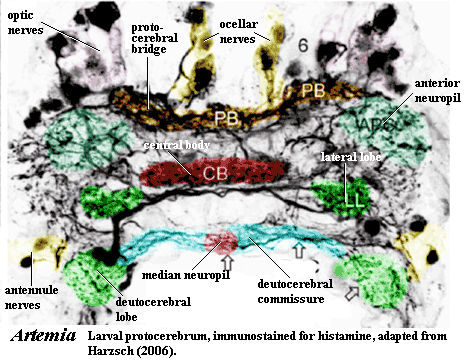 For well over a century, carcinologists have debated whether the most anterior region of the head, the acron, is actually a segment or a pre-segmental region. In part, this is likely a meaningless question. "Segment" is a concept from gross anatomy. It may not always mean the same thing when translated into the languages of developmental genetics, evolution, neurology, etc. Here, we have framed the concept of segment by reference to neuroanatomy. In that sense, the acron is at least a segment, but it may be more.
For well over a century, carcinologists have debated whether the most anterior region of the head, the acron, is actually a segment or a pre-segmental region. In part, this is likely a meaningless question. "Segment" is a concept from gross anatomy. It may not always mean the same thing when translated into the languages of developmental genetics, evolution, neurology, etc. Here, we have framed the concept of segment by reference to neuroanatomy. In that sense, the acron is at least a segment, but it may be more.
The protocerebrum is the neuroanatomical foundation of the acron. Harszch (2006). It seems to have the usual lateral ganglia ("lateral lobes"), apparently connected in some variant of the usual arthropod fashion. However, it also has a large central body which is a median, unpaired neuropil of variable size. The central body seems to be a gigantic version of a small median neuropil, as is sometimes found on the commisures between other ganglia. Note, for example, the small median neuropil on the deutocerebral commisure in the image. In short, the posterior part of the protocerebrum is not all that different from any other segment. For convenience, we'll refer to this as the South End.
Away from the South End, things become more peculiar. Two large anterior neuropils lie anterolateral to the lateral lobes. North of this is the Optic Coast, with two large optic nerves northeast and northwest for the compound eyes, and two small projections medially, for the naupliar eyes. Remarkably, the same basic structure seems to hold, not only crustaceans and insects Harszch, 2006), but also the pycnogonid (sea spider) Anoplodactylus (Maxmen et al., 2005: 1146, fig. 3b) and the millipede Glomeris (Prpic, 2004: fig 2). However, a fully-developed protocerebral bridge is only present in insects and crustaceans. Harszch (2006).
It is tempting to think of the Optic Coast as a sort of half-segment, making the acron a segment-and-a-half. Resist this temptation. That's typological thinking. One reason we suspect the posterior acron is essentially a segment is that the homeobox gene engrailed (en) is expressed along the South End in species which use en to make segments. En is sometimes expressed
CONTINUED ON NEXT PAGE
Page ATW070620
