Panarthropoda: Lobopodia
Taxa on This Page
- HadranaxX
- Orstenotubulus X
- XenusiidaeX
- XenusionX
The Lobopodia or Lobopoda [1] are an evolutionary grade or phylum Cavalier-Smith (1998) of ecdysozoans, intermediate between the worm and the arthropod states. During the Cambrian period, they were very common and diverse, probably filling a range of ecological roles and guilds, although their soft bodies meant that they were rarely preserved, except for lagerstätten and a few rare impressions. This means that their numbers and importance would have been underestimated, as with their contemporaries and distant cousins the /priapozoan worms, another important Cambrian group. If we add the protoarthropods - essentially specialised swimming and gilled lobopodians, this becomes even more the case.
Because the conditions that allowed lagerstätten-type preservation were less common after the Cambrian, it seems that the early lobopodians [2] died out at the end of the Cambrian, but this was not the case, as rare instances have bene found at from the Ordovician and Silurian that hint at a greater diversity ( Whittle et al 2009)
At some point though these early forms disappeared; perhaps a situation similar to the mid-Palaeozoic decline of trilobites and early echinoderms. Eventually, only two specialised lobopodian grade lineages remained, the onychophora, represented today by Peripatus and co, who took up a terrestrial lifestyle, and the tardigrades, enigmatic microscopic forms that independently evolved have many arthropod qualities. All three together are collectively known as "lobopods" or "lobopodians", and they differ from arthropods in not having evolved a hard jointed exoskeleton or efficient, articulated limbs.
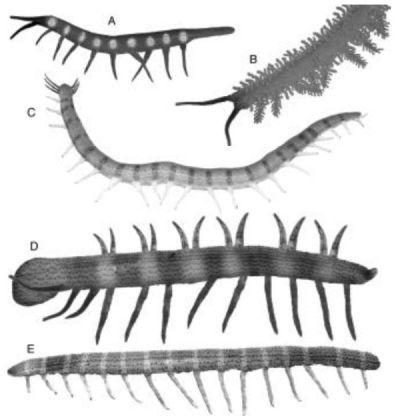 Computer generated reconstructions of the fossil onychophorans. A: Microdictyon sinicum; B: Onychodictyon ferox; C: Cardiodictyon catenulum; D: Hallucigenia fortis; E: Luolishania longicruris. - artwork from Monge-Nájera & Hou (2002) (compare with the line art from Bergstrom and Hou (2001)). These were among the first life-like reconstructions of the variety of Cambriabn lobopodians. Yet while the body proportions and skin texture are probably realistic, Maas et al 2007 have since argued on the basis of the exceptionally preserved three dimensional Orstenotubulus evamuellerae that long-legged lobopodians could not have walked with legs straight such as are shown here (and in numerouis other reconstructions, especially of Hallucinogenia) but would instead have crawled with legs held mostly horizontally. (Ironically Monge-Nájera & Hou in this very paper addressed the problem of taphonomic distortion; the long-legged and straight legged lobopodians would be another example of this). Compare these reconstructions with the more stylised (yet still evocative) images offered by Jerzy Dzik (2011) Computer generated reconstructions of the fossil onychophorans. A: Microdictyon sinicum; B: Onychodictyon ferox; C: Cardiodictyon catenulum; D: Hallucigenia fortis; E: Luolishania longicruris. - artwork from Monge-Nájera & Hou (2002) (compare with the line art from Bergstrom and Hou (2001)). These were among the first life-like reconstructions of the variety of Cambriabn lobopodians. Yet while the body proportions and skin texture are probably realistic, Maas et al 2007 have since argued on the basis of the exceptionally preserved three dimensional Orstenotubulus evamuellerae that long-legged lobopodians could not have walked with legs straight such as are shown here (and in numerouis other reconstructions, especially of Hallucinogenia) but would instead have crawled with legs held mostly horizontally. (Ironically Monge-Nájera & Hou in this very paper addressed the problem of taphonomic distortion; the long-legged and straight legged lobopodians would be another example of this). Compare these reconstructions with the more stylised (yet still evocative) images offered by Jerzy Dzik (2011) |
The onychophores and tardigrades will be described a little later. But it is the Cambrian forms (see illustration, right), such as the venerable Burgess Shale genera Ayshenia and Hallucigenia, as well as Microdictyonand a host of others recovered from the Chengjiang lagerstätte that are the focus of the present unit . Of these Hallucigenia is particulari;ly notorious, due to being spectacularly misinterpreted and reconstructed upside down, leading popular science writer Stephen Jay Gould in his Wonderful Life (on the Cambrian explosion) to wrongly conclude that the Cambrian was full of bizarre short-lived phyla, of which only a tiny perceptage survived. Significantly, several of the organisms Gould described - Hallucigenia, Opabinia, Anomalocaris - are bnow known to form a single evolutionary gradation of transitional forms (see cladograms below).
But while we can now be pretty confident of the big picture, the details elude us. Tardigrades, onychophores, and arthropods all evolved from lobopodians (the latter via protarthropods), but determining exactly which Cambrian lobopodians are related to which remains difficult. Moreover, many Cambriabn lobopodians seem not to be related to modern tardigrades and onychophores at all. A distinction is therefore made between recent onychophores and tardigrades (and their immediate and obvious fossil ancestors) and the range of Cambrian forms. The Class rank taxon Xenusia, which sounds like the name of an alien planet in a 1950s sci fi movie, was coined by Dzik & Krumbiegel, 1989 (from Xenusion) to distinguish the fossil forms from recent onychophores and tardigrades.
With the introduction of cladistic techniques and the study of new taxa, a more developed picture of Cambrian lobopodian morphology has taken shape, beginning with Ramsköld's work on the Chengjiang lagerstätte. In a recent write up, Ramsköld & Chen (1998) analyze the Cambrian lobopodia together with Recent Onychophora, and locate the extant onychophorans as ingroup Cambrian lobopodians on the basis of four synapomorphies. They named two new clades, Alphonychophora and Betonychophora, with extant onychophorans and the Carboniferous Helenodorus making up a third clade, Euonychophora. The three clades constitute an unresolved trichotomy, combined in a monophyletic Onychophora. They reject the possibility of tardigrade relationships. Their cladogram is as follows:
■─┬─Hypanc
└─┬─Betonychophora
│ ├─Paucipodia
│ └─┬─Cardiodictyon
│ └─┬─Microdictyon
│ └─┬─Hallucigenia sparsa
│ └─Hallucigenia fortis
├─Alphonychophora
│ ├─┬─Xenusion
│ │ └─Luolishania
│ └─┬─Aysheaia pedunculata
│ ├─Aysheaia prolata
│ └─Onychodictyon
└─Euonychophora
├─Helenodorus
└─Recent Onychophora
In this cladogram, instead of an outgroup taxon, a hypothetical ancestor (Hypanc) is used. This approach does not seem to be used by many workers.
Budd 1999 however argued that some of the characters considered by Ramsköld and Chen) were invalid or based on outdated concepts such as Articulata (worms + arthropods). For example, "suppression of external segmentation" is not a derived character because lack of external segmentation is a primitive feature shared by Cambrian lobopodians, and ecdysozoa inb general. The features of Cambrian lobopodians are either autapomorphies, such as the long defensive spines of Hallucigenia, or plesiomorphies, such as the annulated trunk and lobopod legs. This is also shown by the fact that proto-anomalocarids such as Kerygmachela and to a lesser extent Pambdelurion, demonstrate the retention of many lobopodian features up to the arthropod stem-group level (Budd 1999, summarised in Budd 2001 and Edgecombe 2009). Nevertehless Ramsköld & Chen (1998) remains an important study, with pertinant observations and critiques.
More recent cladograms depict the Xenusians as a grade of evolution rather than a natural group. The combination of characters revealed by the Cambrian forms provides the opportunity to break up the long branch between recent onychophorans (which are entirely terrestrial) and arthropods (Edgecombe 2009). Nevertheless there is little agreement about actual relationships between the various taxa, as shown by the following series of cladograms
Eriksson et al 2003:
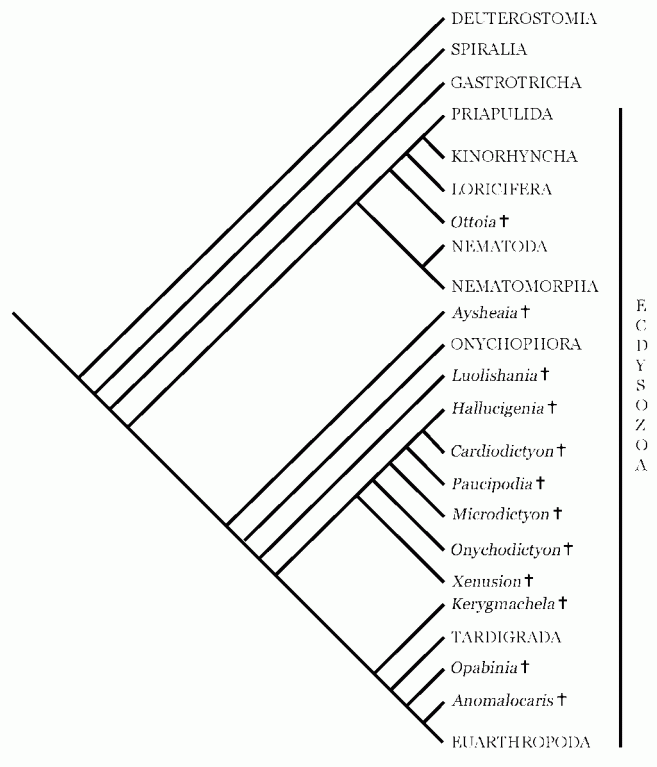
Tree by Eriksson et al 2003 showing possible relationships among extinct and extant ecdysozoans. Extant clades are shown in capitals. Based on Schmidt-Rhaesa et al. (1998), Budd (1996, 1999), and Budd and Jensen (2000). . This innovative cladogram rejects the idea of a monophyletic lobopodia, although most lobopodians still form a single clade. Here Ramsköld & Chen (1998)'s "Betonychophora" constitute the most derived taxa, forming a monophyleticc clade, whilst members of their Alphonychophora make up stem taxa. In keeping with Budd's findings, Ayshenia here occupies a very basal position (Graham Budd, an important and seminal theorist in proto- arthropod development, is co-author of both this paper and the following, see also this earlier mentioned synopsis). In contrast to other phyloggenies shown here, Onychophora is placed beneath (stemward, more primitive than) all other Cambrian lobopodians. The crownward position of the tardigrades shown here (tardigrades as sister taxon of arthropods) is based on Nielsen,1995's influential work, but finds little support in later phylogenetic analyses (Edgecombe 2010).
|
|
Daley et al 2009:
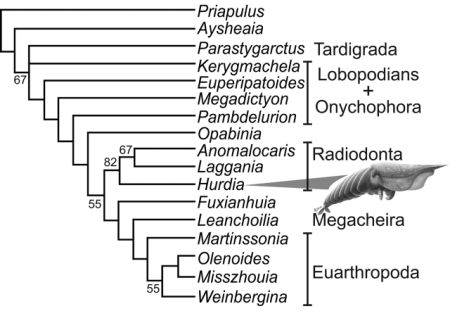
Cladistic analysis of selected stem and crown group arthropods, strict consensus of three trees, from Daley et al 2009's study of the Burgess Shale Anomalocaridid Hurdia victoria (illustrated). An abbreviated version of this cladogram is given by one of the co-authors of this paper, Edgecombe 2009 fig.3 (Although for the sake of convenience only the senior author's name is given, each of the papers featured here has about half a dozen co-authors, more or less). In keeping with the phylogeny provided by Eriksson et al 2003, Ayshenia occupies the basal placement. Several very interesting new taxa put in an appearance here, including the transitional lobopod-protoarthropods Megadictyon and Pambdelurion And although this study mostly features protoarthropods rather than lobopodians proper, it is distinguished by the fact that the lobopod-anomalocarid non-missing link Kerygmachela is here placed even stemward of the onychophora, based on the premise that the ventrally placed mouth (an advanced feature) only evolved once. But in view of the otherwise more advanced features of Kerygmachela, it is more parsimonious to assume that this at least happened twice, once with onychophores (perhaps an adaptation or preadaptation to terrestrial lifestyle) and once with the protoarthropods.
|
|
Ma et al 2009:
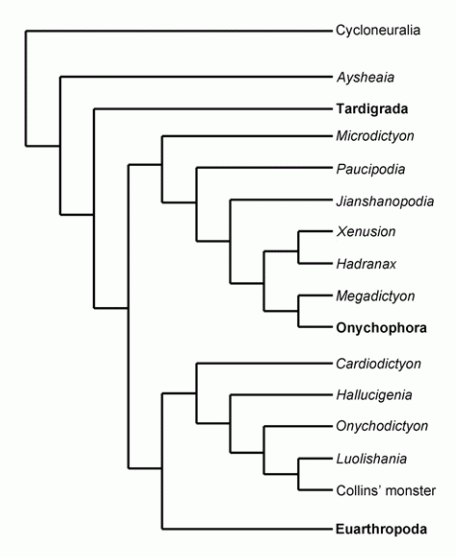 Phylogenetic relationships between Cambrian lobopodians and extant panarthropods from Ma et al 2010, modified from Ma et al 2009. Extant taxa shown in bold. From an analysis of the lobopodian Luolishania longicruris, this cladogram is similar to Ramsköld & Chen 1998 in featuring a large number of lobopodians, and like those authors dividing the lobopodians into several clades, although as with other phylogenies shown here the protoarthropods are also included. Here, apart from Tardigrades and Aysheaia, all lobopodians form two distinct clades. In contrast to Ramsköld & Chen the recent onychophora constitute the crown group of the more stemward of one of these clades (tentatively efferred to as Onychophora). The protoarthropods (anomalocarids etc) and true arthropods constitute sister clades (in contrast to other phylogenies in which they form a single gradation) which is then paired with the more crownward lobopod clade, here called Archonychophora. However, the relationship of the taxa of these two lobopodian clades shows no concordance with either Ramsköld & Chen 1998 or Eriksson et al 2003. The primitive genera Hadranax and Xenusion are also occupy a clade intermediate between the two proto-anomalocaridid-like genera Jianshanopodia and Megadictyon, perhaps due to shared primitive features common to large lobopodians. The sister taxon of the crown group Onychophora is here Megadictyon, which is elsewhere unanimously considered either a basal protoarthropod or a transitional form between lobopodians and protarthropods. Since Euonychophores and Megadictyon are not particularily similar, this makes one think the anomolous result here is an artifact of statistical cladistic method which breaks down when dealing with poor data and numerous homoplasies, as is the case with Cambrian lobopodians. Less controversially, Kerygmachela here returns to a more conventional position at the base of the stem arthropods, above the lobopodians. Phylogenetic relationships between Cambrian lobopodians and extant panarthropods from Ma et al 2010, modified from Ma et al 2009. Extant taxa shown in bold. From an analysis of the lobopodian Luolishania longicruris, this cladogram is similar to Ramsköld & Chen 1998 in featuring a large number of lobopodians, and like those authors dividing the lobopodians into several clades, although as with other phylogenies shown here the protoarthropods are also included. Here, apart from Tardigrades and Aysheaia, all lobopodians form two distinct clades. In contrast to Ramsköld & Chen the recent onychophora constitute the crown group of the more stemward of one of these clades (tentatively efferred to as Onychophora). The protoarthropods (anomalocarids etc) and true arthropods constitute sister clades (in contrast to other phylogenies in which they form a single gradation) which is then paired with the more crownward lobopod clade, here called Archonychophora. However, the relationship of the taxa of these two lobopodian clades shows no concordance with either Ramsköld & Chen 1998 or Eriksson et al 2003. The primitive genera Hadranax and Xenusion are also occupy a clade intermediate between the two proto-anomalocaridid-like genera Jianshanopodia and Megadictyon, perhaps due to shared primitive features common to large lobopodians. The sister taxon of the crown group Onychophora is here Megadictyon, which is elsewhere unanimously considered either a basal protoarthropod or a transitional form between lobopodians and protarthropods. Since Euonychophores and Megadictyon are not particularily similar, this makes one think the anomolous result here is an artifact of statistical cladistic method which breaks down when dealing with poor data and numerous homoplasies, as is the case with Cambrian lobopodians. Less controversially, Kerygmachela here returns to a more conventional position at the base of the stem arthropods, above the lobopodians.
|
|
Liu et al 2011
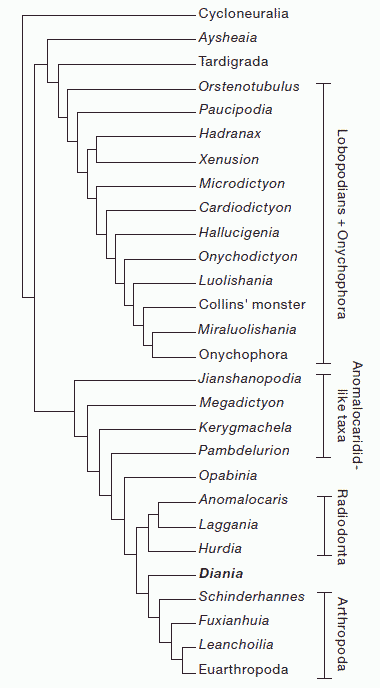 Cladistic analysis of all Cambrian lobopodians and some arthropod stem group taxa, from Liu et al 2011's paper on the "walking cactus" Diania cactiformis, which here resoves as the sister taxon to true arthropods. In contrast to other studies, fossil and recent lobopodians other than Diania here form a single pectinate clade, with extant Onychophora as the most derived member. Ayshenia is the most basal lobopodian, but no longer a basal panarthropod. As with Ma et al 2009, Hadranax and Xenusion are here sister taxa (family Xenusiidae). Ma et al (2009)'s "Archonychophora" is here incorprated with the same order of taxa, except that it is now a paraphyletic group, leading to crown Onychophora. The sister taxon of the Onychophora is Miraluolishania (which Ma et al 2009 had earlier argued was a synonym of Luolishania. , Megadictyon is here returned to the protoarthropods, just beneath Kerygmachela. Ironically, Ma et al (2009)'s sister taxon to the protoarthropods becomes here the sister taxon to the extant onychophora, and vice versa. These strangely inverse placements are clearly the result of lack of obvious synapomorphies, and support Ramsköld & Chen (1998) hypothesis of a number of distinct lineages, and euonychophora not being derived from currently known Cambrian lobopodians. Cladistic analysis of all Cambrian lobopodians and some arthropod stem group taxa, from Liu et al 2011's paper on the "walking cactus" Diania cactiformis, which here resoves as the sister taxon to true arthropods. In contrast to other studies, fossil and recent lobopodians other than Diania here form a single pectinate clade, with extant Onychophora as the most derived member. Ayshenia is the most basal lobopodian, but no longer a basal panarthropod. As with Ma et al 2009, Hadranax and Xenusion are here sister taxa (family Xenusiidae). Ma et al (2009)'s "Archonychophora" is here incorprated with the same order of taxa, except that it is now a paraphyletic group, leading to crown Onychophora. The sister taxon of the Onychophora is Miraluolishania (which Ma et al 2009 had earlier argued was a synonym of Luolishania. , Megadictyon is here returned to the protoarthropods, just beneath Kerygmachela. Ironically, Ma et al (2009)'s sister taxon to the protoarthropods becomes here the sister taxon to the extant onychophora, and vice versa. These strangely inverse placements are clearly the result of lack of obvious synapomorphies, and support Ramsköld & Chen (1998) hypothesis of a number of distinct lineages, and euonychophora not being derived from currently known Cambrian lobopodians.
A new taxon, Orstenotubulus, makes a cladistic debut, it here occupies a position just above the tardigrades, perhaps because of small size. In other respects, Orstenotubulus appears to be a rather typical xenusian lobopodian, with features in common with tardigrades, euonycxhophores, and spiny xenusian lobopodians.
But the most unusal and controversial placement in this cladogram is Diania itself, which apart from its armoured exoskeleton, otherwise resembles a standard vanilla Cambrian lobopodian, is here anomalously placed even above the proto-arthropods. It's elevated position is no doubt due to Liu et al 2011 's premise that arthropodisation and jointed legs only occurred once. But if we assume, like the ventral mouth of onychophores and arthropods, that this occurred several times, Diania would then be located in a more stemward position. Liu et al 2011's analysis was criticised by Mounce & Wills 2011 and Legg 2011, but both critics used a strict consensus approach which resolted in an unresolved polytymy or "star" diagram and which, as Liu et al. 2011b pointed out in their counter-reply, did not provide much meaningful information. In any case, apart from Diania, the protoarthropods and arthropods appear in a pectinate series such as was first proposed by Graham Budd in the late 1990s
|
A characteristic of various lobopodian and protoarthropod cladograms is that whereas there is generally a degree of consensus regarding the latter, there is almost none at all regarding the former. Various taxa are arranged in totally different ways with each analysis. The problem is that lobopodians in general are highly plesiomorphic, and quite likely homplastic, with few obvious distinguishing features, or at least few that are preserved as fossils. Add to that the unreliability of even those characteristics that do appear in fossils. Although lagerstätten (sites of exceptional fossil preservation) such as Chengjiang and the Burgess Shale provide a unique window into the past, by revealing even the soft-bodied organisms and even impressions of internal organs, taphonomic factors inevitably introduce distortion that needs to be considered when reconstructing the anatomy of these ancient craetures. The absence of bucal parts, adhesive-expelling organs, gonopore, eyes, legs, claws, annulation and papillation in fossils may not represent their absence in the actual animals. Leg thickness and claw orientation can also be unreliable, while the apparent presence of certain structures may simply be the result of tissue decomposition (Monge-Nájera & Hou (2002))
An interesting feature of many Cambrian lobopodians, and perhaps a primitive condition for panarthropoda as a whole, are the paired sclerites or minature armour plates,s ymmetrically arranged along the trunk, mostly in pairs dorsally positioned to the walking limbs. These show a general net-like microstructure, which has been described for Hallucigenia, Onychodictyon and Microdictyon, and also found in other genera such as Cardiodictyon, Luolishania (= Miraluolishania?), and "Collins monster"). These may have been a supportive structure for secretive or sensory papillae that may have had tactile, chemosensory or secretory functions, as well as protecting the soft-bodied trunk. Steiner et al 2012 p.121. In support of the lobopodians from worms hypothesis, lobopodian sclerite microstructure is comparable to a similar microstructure in paired trunk sclerites of the (Cricocosmiid palaeoscolecidans Cricocosmia and Tabelliscolex. The net-like trunk sclerites of lobopodians are interpreted as having carried numerous sensory or secretory papillae. Duplicates of trunk sclerites in Microdictyon and Cricocosmia also are evidence of a moulting of the external cuticle. Steiner et al 2012
Some Cambrian lobopodians were well armoured or defended. Hallucigenia sparsa was eqipped with what seem to be long defensive spines. Hou & Bergström 1995 reconstruct Onychodictyon as resembling a stout spiny onychophore. While Diania cactiformis actually did resemble a walking cactus. Jerzy Dzik (2011) reconstructs many lobopodians as prickly customers covered in an array of defensive scleroites, rather like the halwaxiid "coat of mail" animals, and in view of their success in the early Cambrian oceans, it may indeed be that these soft-bodied creatures were well protected against whatever preditors may turn up. MAK120427
Notes:
[1] The taxon Lobopodia was coined by the renowned arthropod morphologist Robert Evans Snodgrass in 1938, to refer to intermediates between annelids and arthropods. The equally brilliant Thomas Cavalier-Smith in 1998 made this the phylum Lobopoda (without the "i"), perhaps to avoid confusion with lobopodia as the technical term for the lobose pseudopods of amoeboid protists.
[2] Grammatically, the unwieldly "lobopodian" is more correct than "lobopod", and hence has bene used here. As explained by Hou & Bergström 2006, a lobopod is the unsegmented, not sclerotised locomotory limb of many panarthropods (or aiolopods to use those authors preferred term), whereas a lobopodian is the animal itself, a panarthropod with lobopod limbs (similarily "pseudopod" is not a taxon but an amoeboid extrusion). For any number of reasons, the term lobopod has taken on the colloquial meaning as shorthand for lobopodian (or lobopodan, or lobopod-bearing animal, depending on your preferred terminology), even appearing in the scientific literature (e.g. Budd 1993 Liu et al 2008). For one thing "lobopodian" can be confusing because the suffix implies it is an adjective (so there would be no way other than context to distinguish it from the noun in the singular). I"Lobopod" has the advantage of being easily and intuitively used, by analogy with arthropod(a) and tetrapod(a), the suffix of the taxon lobopodia in this case is -ia rather than just -a. There is also the convenience, why use five syllables when you can make do with three? With vertebrates by convention we drop the -ia suffix to get the common name, so "Dinosauria" becomes "dinosaur". For now we have chosen to go with the pedantic spelling found in most of the literature, annoying as it may be. Of course there is always the option of adopting Cavalier-Smith's spelling ("lobopoda") to avoid any further grammatical problems and bring lobopod in line with tetrapod, gastropod, and so on MAK120505
Descriptions
Aysheaia pedunculata Walcott 1911
Horizon: Bathyuriscus-Elrathina Zone, Burgess Shale of British Columbia (middle Cambrian). A related species, A. prolata is found in the contemporary Wheeler Formation of Utah
Phylogeny: Panarthropoda : Mureropodia + ((Tardigrada + (Onychophora + (Xenusiidae + Orstenotubulus + Microdictyon + Onychophora + (Paucipodia + Archonychophora + Diania + (paraphyletic Siberiidae + (Kerygmachela + (Dinocaridida + (Schinderhannes + Arthropoda)))))) + *)
Comments: Specimens of Aysheaia pedunculata average about 1 to 6 cm in length, with ten body segments, each of which has a pair of spiked, annulated legs. There are six finger-like projections around the centrally-located mouth and two grasping legs on the "head" (in front of the first pair of trunk appendages). Based on association with sponge remains, it is believed that Aysheaia was a sponge grazer and may have protected itself from predators by seeking refuge within sponge colonies, perhaps using its claws to cling to the sponge (adapted from Wikipedia)
Aysheaia was the first of the Cambrian lobopodians to be recognised as such. It was originally described by Walcott in 1911 as an annelid, along with Canadia (now Hallucinogenia) sparsa. The genus name commemorates a mountain peak named "Ayesha" (originally Aysha) north of the Wapta Glacier, after the heroine of Rider Haggard's 1887 novel She Similarities between Aysheaia and extant onychophora were pointed out to Walcott almost immediately, and Hutchinson in 1930 assigned Aysheaia to its own monotypal order Protonychophora, within the Onychophora (this contains only the monotypal family Aysheaiidae). As a result, and due to overall similarity to the modern day Peripatus, Aysheaia has traditionally been included as an early representative of the modern phylum Onychophora. Whittington (1978) helped popularise the genus in the paleontological community. Other features however, such as cebntrally-located mouth and the ornament of the trunk are distinct from modern onychophorans (Liu et al 2008 p.280),. Simonetta, A. M. & Delle Cave. 1975 pointed out similarities with tardigrades, considering Aysheaia intermediate between Tardigrades and Onychophores (Ramsköld & Chen (1998) p.110). And according to Budd, Aysheaia must be considered a metataxon, as it possesses no currently identified autapomorphies that would distinguish it from the arthropod crown node (Budd 2001 p.274). This is another way of saying that it represents the condition of a common ancestor of that group (in this case panarthropoda). In cladograms by Budd and co-workers, and all later researchers in this field, Aysheaia consistently is located benath the Tardigrade-Onychophore-Arthropod divide, the only exception being Liu et al 2011, for whom Aysheaia is the basalmost member of a monophyletic Lobopodia. (see above cladograms). Featured in many older paleontology books, Aysheaia has since been dethroned by the more charismatic Hallucinogenia, and more recently the range of Chengjiang forms. But the generalised (ancestral) nature of this genus means that it has an important position in the evolutionary history of the panarthropods.
Aysheaia therefore is more primitive then any onychophoran, or even most lobopodians. Such was the rate of evolution at this time that even as early as the Middle Cambrian, Aysheaia wa s a "living fossil", coexisting alongside its more advanced descendants. A parallel may be made with quaternary and recent Monotremes in Australasia, who survive by virtue of specialised morphology and lifestyle. Aysheaia may have had a similar strategy, in this case, as a sponge specialist, it was able to continue in a particular ecological niche long after its contemporaries had disappeared.
We have however placed Aysheaia above Mureropodia because it still is a full-fledged lobopodian, whereas the other two taxon have transitional priapulozoan features. The "half worm" Facivermis, considered a transitional form ( Liu et al 2006), is here reinterprteted as a highly specialised filter feeder.MAK120425
Xenusiidae Dzik & Krumbiegel, 1989
Phylogeny: Panarthropoda : Mureropodia + (Aysheaia + (Tardigrada + ((Paucipodia + Archonychophora + Diania + (paraphyletic Siberiidae + (Kerygmachela + (Dinocaridida + (Schinderhannes + Arthropoda)))) + Orstenotubulus + Microdictyon + Onychophora + * : Hadranax + Xenusion))
Large size, large nodes on the trunk, relatively wide, well-defined trunk annulations, lack of claws Liu et al 2011 although that last feature is likely to simply be due to poor preservation
Comments: large forms so far known only from the Early Cambrian of the baltic and Greenland; may be a small early clade (Liu et al 2011) or a paraphyletic ancestral grade (Dzik 2011). Poor preservation means it is not known whether features such as oral papillae (a feature of modern onychophorans.), a proboscis, or foot claws were present (Liu et al 2011)

Xenusion auerswaldae, impression Kalmarsundzandsteen, found near Sewekow in eastern Germany original url
|
Xenusion auerswaldae Pompeckj 1927
Horizon: Earliest Cambrian (Pleistocene drift), Germany
Phylogeny: Xenusiidae : Hadranax + *
Comments: Although Aysheaia was the first recognised lobopodian (Onychophora sensu latu), Xenusion followed not long after. It was discovered in glacial drift (a sandstone boulder carried by glaciers) in Germany, thought to have originated in Southern Sweden. More recently a second specimen was discovered, and described by Dzik & Krumbiegel, 1989. Both specimens were preserved as empty cuticles shed after moulting. Originally considered to be of Late Precambrian (Late Neoproterpozoic) age, Xenusion is now considered to be Earliest Cambrian. It was a large lobopodian with pairs of dorsal plates; From Wikipedia: "The specimens are not especially well preserved. The older specimen is 10 cm or so in length with a narrow, weakly segmented body. A depression runs up the bottom on all but the rearmost segments. There is a slightly bulbous tail, and each segment beyond that seems to have a single pair of tapering annulated legs similar to the modern onychophore, but without claws. Nine segments are present. There is a spine on each body bump and faint transverse parallel striations on the annulations on the legs. The legs of what is possibly the foremost segments are either missing or not preserved. The head is believed to be missing or is poorly preserved. " Although Xenusion was armed with spines, they were shorter than those of Hallucigenia. The original specimen would appear to be part of an animal about 20 cm in length, larger than most Cambrian species or extant onychophores.
Considered an ancestral form intermediate between Tardigrades and Onychophores by Dzik, who places it at the base of his dendrogram, largely also because of the date (earliest known lobopodian) (Dzik & Krumbiegel, 1989, Dzik 2011). Ramsköld & Chen (1998) argue that Dzik & Krumbiegel (1989)'s interpretation of the second specimen is incorrect, in that it should be interpreted as preserved from the ventral (bottom) rather than the dorsal (top) side. They also state that there is no evidence of a long, (priapulozoan-like) proboscis (although this does not mean it would have been absent, simply that it wasn't preserved), nor is the front of the trunk narrower than the rear.
The phylogenetic status of this taxon are debatable, as typical with the Xenusia as a whole, and so far cladistic analyses has failed to resolve the problem. Ramsköld & Chen 1998 include it in their Alphonychophora, along with Luolishania, Ayshenia and Onychodictyon. Dzik 2011 place it close to Aysheaia but not to the other two taxa. Eriksson et al 2003 consider it a stem-xenusian (in an unnamed clade), but more derived than recent onychophora. While according to both Ma et al 2009 and Liu et al 2011 it is a stem-onychophora, more derived than basal taxa such as Paucipodia but still fairly primitive in relation to recent onychophora (i.e. it it tends to resolve at about halfway up the cladogram). In view of all this uncertainty we have for now retained it as a general basal lobopodian. MAK120501.
References: Dzik & Krumbiegel, 1989; Ramsköld & Chen (1998)
Hadranax augustus Budd and Peel, 1998
Horizon: Early Cambrian Sirius Passet lagerstätte, North Greenland
Phylogeny: Xenusiidae : Xenusion + *
Comments: A large lobopodian known from an incomplete specimen, it has strong appendages and dorsal tubercles; it seems to be related to Xenusion (Budd & Peel 1998)
 Orstenotubulus evamuellerae, the only lobopodian preserved in three dimensions (apart from Cambrian tardigrades which however are more derived forms). The exceptional preservation, down to the microscopic scale, make this the best understood of all the Cambrian armoured lobopodians. Photo © CORE Orsten Research. This photograph does not show the large spines (see sketch below left)
Orstenotubulus evamuellerae, the only lobopodian preserved in three dimensions (apart from Cambrian tardigrades which however are more derived forms). The exceptional preservation, down to the microscopic scale, make this the best understood of all the Cambrian armoured lobopodians. Photo © CORE Orsten Research. This photograph does not show the large spines (see sketch below left) |
Orstenotubulus evamuellerae Maas et al 2007
Horizon: "Orsten" beds, Västergötland, Sweden, Agnostus pisiformis Biozone (formerly zone 1 of the Upper Cambrian succession in Sweden).
Phylogeny: Panarthropoda : Mureropodia + (Aysheaia + (Tardigrada + ((Paucipodia + Archonychophora + Diania + (paraphyletic Siberiidae + (Kerygmachela + (Dinocaridida + (Schinderhannes + Arthropoda))))) + Xenusiidae + Microdictyon + Onychophora + *))
Comments: The first Cambrian lobopodian preserved in three dimensions, the exquisate micro-preservation of Orstenotubulus has provided many important insights about the early history and morphology of this group. Fossils are extracted by acid preparation and, as with all the orsten fossils, provide a degree of preservation down to the level of microscopic detail, providing an exception opportunity for study of the morphology of these ancient invertebrates.
All the Orsten fossils are tiny, on the millimeter scale or less. At 4 to 5 mm in length, and a diameter of about 100 to 200 microns, Orstenotubulus is only a tenth the linear dimensions of the better-known Cambrian lobopodians, although in oall ther respects very similar. There are 9 or 10 leg-bearing segments, and body and limbs are almost totally cylindrical. Orstenotubulus not only reveals the unscleratised, micro-annulated body and unjointed annulated limbs (shared with other Cambrian and recent lobopodians). The pattern of micro-annulated regions alternating with smooth regions also occurs in the Chengjiang lobopodians Cardiodictyon and Microdictyon. The micro-annulation indicates that the body of Orstenotubulus was only flexible in the regions between the limbs, whereas the smooth regions were most likely sclerotized. These give rise to tubular legs that extend laterally. In some segments, the smooth zones are dorsally drawn out into paired, conical humps or domes, extending into long spines. Such paired spines arranged in rows on conical humps or domes are characteristic of several Cambrian lobopodians, such as Xenusion, Hallucigenia, and Luolishania, possibly protecting the animal from predatorsand presumably possessing telescopic movability. Hallucigenia sparsa even had similar domes. Comparable dorsal outgrowths and telescopic spines occur in some Tardigrada but are absent in extant Onychophora, The legs are also equipped with spine-like outgrowths occur on short, conical sockets. These were probably telescopic and retractable. Similar features seem to occur in Aysheaia, Xenusion, Luolishania, and Jianshanopodia , but tardigrades and onychophorans lack such structures.
Towards the ventral midline of each leg, the spine-like outgrowths are accompanied by sets of outgrowths, or dermal papillae, consisting of several epidermal cells. They may also have had apical bristles (their absence here may be a preservational or preparational artifact), are similar to tubercles scattered over the body surface of Aysheaia, as well as to those of extant onychophora. These papillae might have had a sensory function, such as coordinating the retraction of spines during locomotion, and be an ancestral characteristic for panarthropods as a whole.
O. evamuellerae also shows strikingly similarities with recent tardigrades and onychophorans in the surface microstructure of its cellular-structured cuticle and the telescopic spines. The cuticle is annulated with a hexagonal surface microtexture like that of extant onychophorans.
The single ventral gonopore (genital opening), which indicates that this feature, also found in onychophorans, tardigrades, pentastomids, myriapods and insects, would seem to represent the plesiomorphic state for the panarthropoda. The paired state in chelicerates and crustaceans was then convergently achieved. (As with the general body plan and the plesiomorphic embryology of uniramia as opposed to more derived arthropoids, this shows that these lineages predate more specialised forms. Hence it is unlikely that the Pancrustacea hypothesis (insects evolved from Crustaceans) is correct)

Orstenotubulus is, like that of many other Cambrian forms, of the long-legged type, a morphotype with no extant equivalent. The specimens demonstrate that "ventral dangling" might not even occur after death, and its frequent occurance in other fossil lobopodians is the result of distortion due to two dimensional preservation, such as the animal being covered by sediment and its leg pairs pressed together, although lateral posture might be true of at least some short-legged forms, such as Aysheaia and Jianshanopodia. Hence walking on long, unsclerotized unjointed limbs, such as current reconstructions of Hallucinoghenia and Micrtodictyon, is not possible. This shows how misleading it can be to rely only on extant forms, such as onychophores, tardigrada, and arthropods, where changes in locomotory mechanisms have already taken place. The lateral orientation of Orstenotubulus' limbs. supported by anchoring spines pointing ventrolaterally, imply that most early lobopodians were crawlers rather than walkers MAK120427
Reference: the above is condensed from Maas et al 2007 and a few second-hand sources,a long with our own commenst ina few places.
Link (and images): CORE Orsten Research

