Ecdysozoa: Panarthropoda
Taxa on This Page
- Mureropodia X
- Panarthropoda
During the Cambrian period, ancestral Panarthropods were much more diverse than they were even during the later Paleozoic. There were several different lineages of lobopodians and proto-arthropods, including large carnivorous swimming forms like Anomalocaris and Kerygmachela, as well as smaller crawling types such as Aysheaia, Hallucigenia, and Microdictyon, and larger crawlers such Xenusion , Jianshanopodia, and Megadictyon. Whilst true arthropods, tardigrades and onychophorans are highly differentiated in form today, such was not always the case. During Cambrian Epoch 2, many transitional forms co-existed, and exciting new discoveries have come to light. Of particular interest are froms such as the aforementioned Jianshanopodia, Megadictyon, and Kerygmachela, which provide a link between lobopodians on the one hand, and anomalocaridids and arthropods. on the other. Another new anomalocarid-like taxon, Pambdelurion , indicates that the biramous arthropods actually arose from within the anomalocaridids (Budd 1997). After dominating the oceans for many millions of years, the big protoarthropods seem to have disappeared by the Mid Ordovician, their place as super-predators being taken by large cephalopod mollusks. However, medium-sized forms (still respectable in invertebrate standards) such as Schinderhannes continued to the Devonian at least. MAK020503, revised ATW050113 and MAK120505
 Graham Budd's scenario of panarthropod evolution, showing successive morphological innovations, and correlated functional and ecological shifts, with particular reference to feeding mode, from Budd 2003. Larger diagram. A version of this diagram (modified from Budd, 1996, 1999), is also at Species splitting)
Graham Budd's scenario of panarthropod evolution, showing successive morphological innovations, and correlated functional and ecological shifts, with particular reference to feeding mode, from Budd 2003. Larger diagram. A version of this diagram (modified from Budd, 1996, 1999), is also at Species splitting) |
Dzik and Krumbiegel (1989) hypothesize an origin from priapulid-palaeoscolecid-like worms through the development of segments and limbs. Complete specimens of Facivermis yunnanicus, showing both priapulid and arthropod characters, provide strong support for this proposed affinity. (ref Cambrian lobopods), although this could be taken the other way as well, Facivermis being a lobopodian in the process of losing its limbs (see also the discussion on worms and lobopodians).. We have followed Dzik 2011 in making Facivermis a specialised end-taxon far from the original ancestral lobopodian condition. Better evidence may be had in the discovery of a new, priapulozoan-like lobopodian, Mureropodia apae
The bauplan of early panarthropods (paraphyletic class or phylum Xenusia) includes a vermiform body; a proboscis or mouth cone; paired lobopods with claws; a cuticle displaying a repeated anatomical patterning; a straight digestive tract with terminal mouth and anus (Vintaned Gamez et al 2011). The early panarthropods quickly evolved through a series of stages of progressive arthropodisation, such as are shown in this diagram, based on the work of formost specialist and theorist in proto-arthropd evolution, Professor Graham Budd.. The actual sequence is probably not as clear cut and linear as is shown here, because of mosaic evolution and the fact that many traits would have evolved differently and in parallel, e.g. the otherwise primitive lobopodian Diania evolved a sclerotised exoskeleton before seqmentation, compound eyes, biramous limbs and so on. In this context, Cope and Osborne's idea of orthogenesis seems to have some merit, albeit more as parallel adaptive trends of mosaic evolution, as the arthropod potential would already be there in the lobopodians. The degree of homoplasy and parallelism is probably the reason why it is hard to get a consistent phylogeny; every worker in the feld seems to arrive at a different toplogy. In more recent trees for example, Pambdelurion is consistently less derived, rather than more derived, than Opabinia (e.g. Edgecombe 2009, Daley et al 2009, Liu et al 2011)
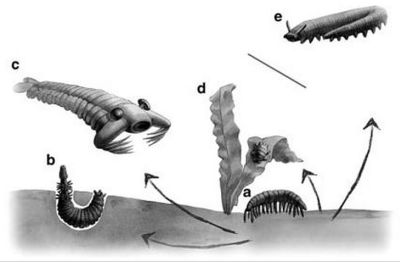 Generalised reconstructions of early Cambrian xenusian (a), cephalorhynchian worm (b), anomalocaridid (c), tardigrade (d), and extant terrestrial onychophoran (e), according to the phylogeny proposed by Vintaned Gamez et al 2011. Artwork by Anastasia Besedina. See also Zhuravlev et al 2011 fig 5 for a more detailed version of this scenario. Generalised reconstructions of early Cambrian xenusian (a), cephalorhynchian worm (b), anomalocaridid (c), tardigrade (d), and extant terrestrial onychophoran (e), according to the phylogeny proposed by Vintaned Gamez et al 2011. Artwork by Anastasia Besedina. See also Zhuravlev et al 2011 fig 5 for a more detailed version of this scenario. |
Vintaned Gamez et al 2011 and Zhuravlev et al 2011 suggest that morphologically diverse xenusians (basal lobopodia) could have given rise to four different ecomorphic lineages: to cephalorhynchian or cycloneurian worms by adaptation for burrowing with retractable proboscis, via forms such as Facivermis and Mureropodia; to tardigrades by adaptation for interstitial habitat (via Hadranax-like forms); to euarthropods by adaptation to walking on joint appendages (via Jianshanopodia like forms); and to anomalocaridids by adaptation to swimming with lateral flaps (via Kerygmachela and similar types). This scneario is shown in the diagram at the left. We believe however that it is more likely that priapulozoan worms came first, as per Dzik and Krumbiegel 1989, Budd 2001, Conway Morris & Peel 2010, etc, and that early lobopodians were transitional forms between these and tardigrades, onychophores, and protoarthropods (anomalocaridids etc) MAK120422
Descriptions
Panarthropoda Nielsen, 1995 = Haemopoda Cavalier-Smith 1998 = Aiolopoda Hou and Bergström, 2006 (lobopodians (note) + arthropods)
Phylogeny: Ecdysozoa : Scalidophora + (Nematoida + (Cricocosmiidae + * : Mureropodia + (Aysheaia + (Tardigrada + (Xenusiidae + Orstenotubulus + Microdictyon + Onychophora + (Paucipodia + Archonychophora + Diania + (paraphyletic Siberiidae + (Kerygmachela + (Dinocaridida + (Schinderhannes + Arthropoda)))))))))
body segmented, with limbs on several segments; adult body cavity a haemocoel that extends into the limbs Cavalier-Smith (1998), "cuticle of a-chitin which is molted regularly, appendages with chitinous claws, and mixocoel with metanephridia and ostiate heart (absent in tardigrades); several characters, such as the structure of the brain, indicate that arthropods and tardigrades are sister groups." Nielsen, 1998, paired segmental ventrolateral appendages, 2 paired leg nerves, Engrailed gene expressed in posterior ectoderm of each segment (Gabriel and Goldstein 2007, cited in Edgecombe 2009). Primitively possess a terminal mouth, a non-retractable proboscis, and a thick integument of diverse plates. Zhuravlev et al 2011, A single differentiated cephalic appendage appears to be a synapomorphy of onychophorans and euarthropods Budd 2001
Comments: [1] Panarthropods include true arthropods and their soft-bodied relatives, the tardigrades and onychophorans. They are generally considered a monophyletic taxon, although there is still some uncertainty regarding the phylogenetic status of the tardigrades, this is more likely to be an artifact of molecular analysis rather than reflecting actual phylogeny (Rota-Stabelli et al 2010), an alternative position that cycloneuralian worms are derived (i.e., evolved from) lobopodians has minority support (Vintaned Gamez et al 2011, Zhuravlev et al 2011) but is not widely held.
[2] The Lobopodia constitute a paraphyletic assemblage that includes all non-euarthropodous panarthropods. It is considered a phylum (with alternate spelling, no "i") by Cavalier-Smith (1998). MAK120425
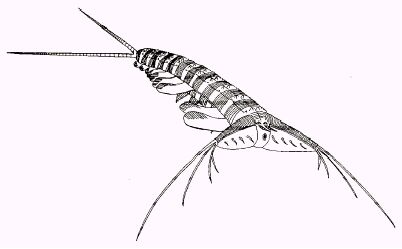
Image: The proto-arthropod Kerygmachela kierkegaardi Buen Formation, Cambrian Epoch 2, North Greenland. From the Transactions of the Royal Society of Edinburgh: Earth Sciences, volume 89 (1999 for 1998), pp. 249290.
Mureropodia apae Vintaned Gamez et al 2011
Horizon: Murero Lagerstätte, bBase of Level RV 1/5 (Protolenus jiliocanus zone, upper Bilbilian, Latest Early Cambrian) in the Cadenas Ibéricas, NE Spain
Phylogeny: Panarthropoda : (Aysheaia + (Tardigrada + (various lobopodia + Onychophora + (Archonychophora + (paraphyletic Siberiidae + (Kerygmachela + (Dinocaridida + (Schinderhannes + Arthropoda)))))))) + *
Comments: Mureropodia apae may well represent a non-missing link between priapozoa and lobodous panarthropods, in which case the dendrograms on these pages need to be revised accordingly. The skin has a fine regular reticulated pattern and is covered by tiny wart-like protuberances and plates, and may have had hair-like sensorial organs as tactile and chemoreceptors. It had at least five pairs of short legs equiped with claws, that may have been used to burrow in the seabed. The length/body width ratio reveals that this animal hardly was able to walk on the bottom surface, and more likely used the limbs for anchoring the body to the substrate. There was a well-developed muscular system and a long retractile proboscis for catching prey. The exceptional fossil preservation has enabled researchers describe interwoven muscle fibres distributed throughout the body, and a dermomuscular sac. Although Mureropodia may have been able to crawl along the surface, it was better adapted to an infaunal, endobenthic lifestyle, burrowing by peristaltic movements.
Reference Vintaned Gamez et al 2011
Links Descubierto un enigmático fósil ancestro de los artrópodos Investigadores valencianos descubren un fósil de hace 520 millones de años (Spanish), Mureropodia, worm-arthropod link? (brief English comment on previous), Proyecto Murero MAK120422
Images: from first two links above
 Graham Budd's scenario of panarthropod evolution, showing successive morphological innovations, and correlated functional and ecological shifts, with particular reference to feeding mode, from Budd 2003. Larger diagram. A version of this diagram (modified from Budd, 1996, 1999), is also at Species splitting)
Graham Budd's scenario of panarthropod evolution, showing successive morphological innovations, and correlated functional and ecological shifts, with particular reference to feeding mode, from Budd 2003. Larger diagram. A version of this diagram (modified from Budd, 1996, 1999), is also at Species splitting)
