Brachiopoda
 Apart perhaps from the trilobite, no other organism typifies the Age of Invertebrates more than the brachiopod. They were the first bilaterian animals to lose their mobility and encase their bodies in a solid external shell. Despite a superficially similar appearance to clams, they are actually completely different in their anatomy, belonging to the group known as the Lophophorata, which use a fringe of tentacles known as the lophophore to sweep food particles into their mouths. Brachiopods are rare today, but during the Paleozoic era especially from the Middle Ordovician period onwards) they absolutely dominated every benthic (bottom-living) marine ecology, their shells accumulating in countless billions. Today their petrified remains are the most common of all fossils.
Apart perhaps from the trilobite, no other organism typifies the Age of Invertebrates more than the brachiopod. They were the first bilaterian animals to lose their mobility and encase their bodies in a solid external shell. Despite a superficially similar appearance to clams, they are actually completely different in their anatomy, belonging to the group known as the Lophophorata, which use a fringe of tentacles known as the lophophore to sweep food particles into their mouths. Brachiopods are rare today, but during the Paleozoic era especially from the Middle Ordovician period onwards) they absolutely dominated every benthic (bottom-living) marine ecology, their shells accumulating in countless billions. Today their petrified remains are the most common of all fossils.
Brachiopods are marine shelled invertebrates that look superficially very much like clams. They are actually quite different from clams in their anatomy, and they are not closely related to mollusks at all. In fact they belong to a (possibly polyphyletic) group known as the Lophophorata, because they feed by using a fringe of tentacles known as the lophophore. These tentacles sweep microscopic food particles in the water into the creature's mouth. Most species of brachiopods are attached to the substrate by a muscular stalk, known as the pedicle. There is however still a free-floating larval stage.
Sixteen hundred genera and many more species of Brachiopoda are known altogether, the vast majority being fossil (extinct) forms.
The Brachiopod Shell
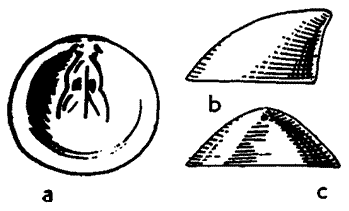
Brachiopods superficially resemble bivalve mollusks in that the animal secretes a bivalved (two-part) shell of calcium carbonate or a combination of calcium phosphate and chitinous organic substance. However, Bivalve mollusks generally have shells that are equal in size and shape (although mirror images of each other), whereas the two shells of brachiopods are of unequal size (the technical term is inequalvalved). The valve (shell) that has the attachment for the pedicle is the pedicle valve which is usually the lower and larger valve. This valve includes the pedicle opening.
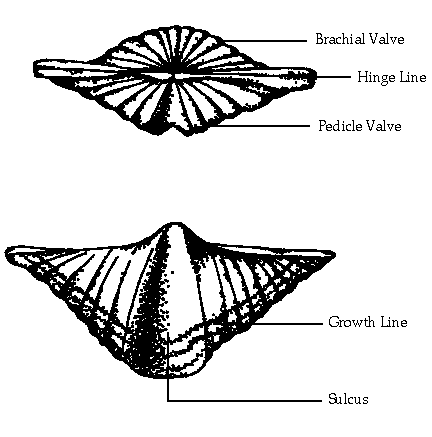
The valve that holds the a feeding tentacles - the lophophore or brachia - is called, naturally enough, the brachial valve. The brachidium, a long ribbon or loop-shaped calcified support for the lophophore, is attached to this shell. The brachial valve is usually, but not always, on the dorsal ("top") side of the organism.
Usually on the pedicle valve there is a raised area called the fold with a complimentary depression, the sulcus, on the brachial valve.
The area where the two valves are in permanent contact is the hinge. The hinge may extend across the entire posterior of both valves, in which case the shell is called Strophic. In Non-Strophic brachiopods the hinge axis passes through teeth and sockets and these serve as a fulcrum for the opening and closing of the shell.
The brachiopods have traditionally been divided into two classes, the Inarticulata and the Articulata. This classification is now no longer considered valid by many workers in this field. Nevertheless it is still useful as a functional analysis. In articulate brachiopods the hinge axis is lined with a set of interlocking teeth and sockets. Inarticulate brachiopods, as their name indicates, lack these teeth and sockets by which to articulate.
Anatomy

The interior of the shell is lined with a mantle, a membranous duplication of the body wall, through which respiration may occur and which secretes the shells. The shell is closed and opened by adductor and diductor muscles respectively. The scars of these muscles may be seen on the inside of the valves. The body of the animal occupies only about one-third of the interior of the shell. The rest is taken up by the lophophore, which is supported by a two-limbed, calcareous structure, the brachidium. The brachidium is variable in shape consisting in its simplest form of the loop of two short or moderately long, curved structures, and in its more complex form of two thin, spirally coiled ribbons or spires. The shape of the brachidium is very important in determining the classification of brachiopod types. The brachidium supports the brachia, the fleshy arm-like part of the lophophore which bear the tentacles or cirri that sweep food particles into the mouth.
Ecology
Unlike their cousins the bryozoans, brachiopods are solitary, and never form colonies. They are limited to marine environments, although some inarticulate brachiopods like Lingula tolerate brackish water. During the early to mid Paleozoic Lingulids lived in association with armoured jawless fish (Ostracoderms) in marginal marine environments.
Brachiopods are sessile (attached to a substrate), benthic (bottom-living) suspension-feeders, feeding on micro-organisms or organic particles dissolved in the water. Most brachiopods were or are epifaunal, living on the surface of the sea-bottom, but a few, like the very primitive and ancient inarticulate genus Lingula is infaunal, burrowing in the sediment. Some articulate species also apparently lived partially buried in the sediment, but these were exceptional. Generally brachiopods differed from bivalves in that bivalves have explored and taken over the infaunal (buried) zone, something the articulate brachiopods were (with a few exceptions) unable to do.
Recent species attached to a surface by the pedicle. Some species have pedicles that are fibrous and muscular and can elevate and rotate the shell directly, while in others the pedicles are inert anchors. Some extinct brachiopods either have no pedicle openings or have a tiny opening for a pedicle that served a tethering function, but did not support the shell. These species were free lying. These did not attach to the substrate but simply grew in piles. Some of these species had a functional pedicle in the juvenile stage, but became free-lying as adults. Going to the other extreme some recent inarticulates and some fossil articulates, cemented their shells to the substrate.
Brachiopods have evolved to use very little energy or oxygen. Consequently they are found in marginal environments such as deep-water, dysaerobic and brackish settings. In the past brachiopods were often a dominant organism in their ecosystem. Usually several species are found together.
Benthic assemblages of brachiopods are correlated with increasing depth. The determining factors were temperature, water pressure, water turbulence, salinity, substrate and food supply.
Evolutionary History
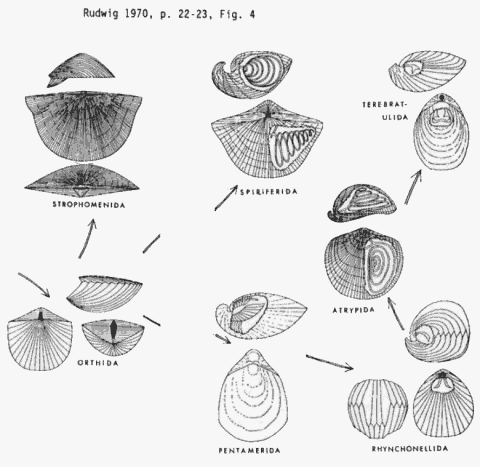
Despite their unassuming character, the brachiopods constitute one of the most important and abundant groups of Paleozoic marine organisms. They diversified into a number of different morphologies and even participated in the build-up of ancient reefs. Certainly in terms of numbers of fossil remains they are the most numerous of any paleozoic organism. Fossil brachiopods are a favourite subject for paleontologists because of their abundance, diversity, and usefulness in stratigraphic correlation. They are much less important in neontological zoology, and most of the knowledge of modern species is based upon old studies.
Brachiopods first appear at the very beginning of the Cambrian Period. During this time they were almost entirely (apart from a few rare types) Inarticulates. There were a number of lineages of Inarticulate type. These developed either or both the strophic (wide) hinge and the calcitic shell that typify the articulate condition. The first true articulates, the strophic order Orthida also appear at this time. The syntrophidiine (strophic hinged) pentamerids appeared a little later, during the Middle Cambrian. Some of the small Inarticulate orders died out about this time.
The Articulate Brachiopods (see above diagram for representative types) expanded greatly in numbers and diversity during the Ordovician. It was during this period that they attained a pre-eminence that they would hold until the end of the Paleozoic era. 200 genera are known from the Late Ordovician alone, but this is probably only a tiny fraction of the real number of types alive then.
The Early Ordovician saw not only a great increase in the Orthids and Pentamerids, but also the origin of two important orders, the strophomenids and the rhynchonellids. The latter represented the first truly non-strophic (narrow hinge) shells with a purely internal articulation (teeth-sockets); a configuration more like that of bivalves. The rhynchonellids in their turn seem to have given rise to the atrypids and athyrids spire-bearers. It is now believed that these types evolved a spiral or coiled lophophore independently of true spirifids, which appeared during the Late Ordovician-.
The Ordovician represented the heyday for the Orthids, which outnumbered every other order of brachiopod. The Inarticulates also seem to have reached their peak around this time, although they were rather rarer than the Articulates.
The terminal Ordovician extinction hit the Orthids badly. By the start of the Silurian they were no more diverse than the other Brachiopod lineages. The Silurian in general seems to have been a rather uneventful time for brachiopod evolution. At this time the Inarticulate types had become quite rare, and they were never again to retain their Cambrian status.
During the Devonian period the articulate brachiopods underwent another great evolutionary radiation, like that of the Ordovician. This period represented the peak of diversity for the Spiriferida. So abundant were these brachiopods that the Devonian period can well be called the "age of spiriferids". At the same time important new groups like the strophomenids and the terebratulids appeared. Conversely there was a general decline and widespread extinction of pentamerids and atrypid spiriferids.
The Carboniferous period saw the Spiriferids supplanted by the strophomenids, especially the productids, which were to remain the most important brachiopod group until the end of the Permian. During this time the other orders of brachiopods declined slightly (e.g.Spiriferids, Orthids), or remained at the same stage of diversity as they had attained during the Devonian.
The Brachiopods were much reduced by the terminal Permian extinction, with the Orthids being wiped out entirely. The spiriferids and strophomenids made it into the Triassic in reduced numbers, the strophomenids being now very rare. Most of the remaining Spiriferids were knocked out by the Triassic terminal extinction, with only a few stragglers making it through to the Jurassic.
The Terebratulids and Rhynchonellids staged a mild resurgence during the Jurassic, while during the same period the spiriferids and strophomenids died out completely.
By the Cretaceous period only the Terebratulids and Rhynchonellids remained in any number. They were accompanied by two surviving orders of Inarticulates and the enigmatic Thecideida.
The Brachiopods now make up a relatively insignificant element in most marine faunas. Most surviving forms are found in parts of the Pacific Ocean, mostly in very cold water, either in polar regions or at great depths in the ocean. There are seventy surviving genera and about 300 living species of brachiopods.
Inarticulate brachiopods
Articulate brachiopods
Links
University of California Museum of Paleontology -- Introduction to the Brachiopoda
Brachio-Philes
- a good up to date intro to the Brachiopods
The Centre for Recent Brachiopod Taxonomy
BrachNet - this European Web site includes up-to-date information on Brachiopoda, references, systematics, diagnoses...) a directory of specialists working on brachiopods (Brachiopodologists - yes, there is such a term!).
References
Sokiran, EV (2002), Frasnian–Famennian extinction and recovery of rhynchonellid brachiopods from the East European Platform. Acta Pal. Pol. 47: 339–354.

Brachiopoda main page
|