 Our working phylogeny of the Deuterostomia is discussed below. In outline, it looks like this:
Our working phylogeny of the Deuterostomia is discussed below. In outline, it looks like this:| Deuterostomia | ||
| Deuterostomia | Deuterostomia |
| Metazoa | ||||||
| Metazoa | Metazoa | Metazoa | Time |
|
Metazoa
├─Porifera (paraphyletic?)
└─┬─Radiata
└─Bilateria
├─Deuterostomia
│ ├─Vetulicolia
│ └─┬─Chordata
│ └─Ambulacraria
│ ├─Hemichordata
│ └─┬─Vetulocystidae
│ └─Homalozoa
│ └─Echinodermata
└─Protostomia
├─Ecdysozoa
└─Lophotrochozoa
|
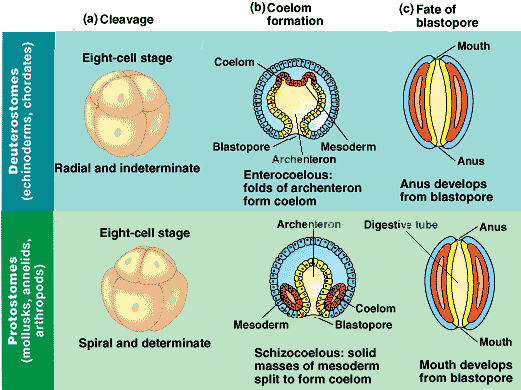
Compared to the protostomes, the deuterostomes are far less diverse, including among the invertebrates only the Echinoderms and a few minor taxa, but having as their main claim to fame the fact that Chordata belong to this sub-division. The deuterostome condition is often associated with an initial embyonic cleavage pattern which is "radial" and in which cleavage planes are either parallel or perpendicular to the vertical axis of the embryo. Deuterostomes also commonly exhibit an indeterminate cleavage in the early stages of embryo development. That is, each early embryonic cell retains the capacity to develop into a complete embryo if isolated from other cells. (this enables the production of twins in vertebrates.). Deuterostomes also possess enterocoelous coelom development, in which the mesoderm arises as lateral outpocketings of the archenteron (embryonic gut) with hollows that become the coelomic cavities, and the development of the embryonic blastopore (the original opening) into the anus (rather than into the mouth as in most protostomes). MAK020407, rev'd ATW050814.
Our working phylogeny of the Deuterostomia is discussed below. In outline, it looks like this:
 The deuterostomes make up on of the earliest divisions with the Bilateria. Included in this clade are three very different-looking phyla - the echinoderms, hemichordates and chordates.
The deuterostomes make up on of the earliest divisions with the Bilateria. Included in this clade are three very different-looking phyla - the echinoderms, hemichordates and chordates.
The characters uniting these taxa are mostly developmental (Miller & Harley, 1996). The name Deuterostomia ("second mouth") refers to the development of the embryo blastopore into the anus, with the mouth developing as a secondary opening, in contrast to protostomes ("first mouth") with the process reversed. The initial embryonic cleavage in deuterostomes is radial and indeterminate, and coelom development is from outpockets of the gut (enterocoelous). Molecular data have supported monophyly of the Deuterostomia.
This section will cover the deuterostomes that lie outside the echinoderm and craniate crown groups. Deuterostomia is here defined as all animals more closely related to Homo Chordata) and Echinus (Echinodermata) than to Scarabaeus (Arthropoda) or Helix (Mollusca). It should be noted that "deuterostomy" (the anus developmentally proceeding the mouth) is not unique to Deuterostomia, but is also found in Bryozoa, Brachiozoa and Chaetognatha. On this page, "Deuterostomia" and "deuterostome" always refer to the clade, while "deuterostomy" and "deuterostomous" refer to the developmental pathway.
The evolutionary relationships of deuterostomes have gone through a fair amount of contention in recent years. Traditionally. mainly after the work of Hyman, deuterostomes have been placed quite high up in the animal tree, forming a clade with the other deuterostomous taxa nestled within the coelomate clade see Bilateralia 2). Within the Deuterostomia, chordates and hemichordates were united by possession of ciliated pharyngeal gill pores/slits and a stiffened notochord-like structure (the notochord in chordates, the stomochord in hemichordates). The similarities in larval form between hemichordates and echinoderms were regarded as ancestral for deuterostomes, and subsequently lost in chordates.
 Humans being the chauvanistic animals we are, there's a lot of appeal in this tree, which appears to sit us at the top of ever-increasing complexity. Unfortunately, recent phylogenies, mostly molecular, rather put the kybosh on this view (Maddison, 2002). Deuterostomia is much lower down, probably the second branch of Bilateria to split off after the Acoelomorpha. Compared to its sister group, the omnipresent and spectacularly diverse Protostomia (containing the arthropods, molluscs and nematodes as its particular success stories), the deuterostomes are merely minor players. Interestingly, while protostomy has been generally regarded as ancestral to deuterostomy, modern phylogenies suggest the opposite, as shown by this phylogeny based on Giribet et al. (2000) where D = deuterostomous and P = protostomous:
Humans being the chauvanistic animals we are, there's a lot of appeal in this tree, which appears to sit us at the top of ever-increasing complexity. Unfortunately, recent phylogenies, mostly molecular, rather put the kybosh on this view (Maddison, 2002). Deuterostomia is much lower down, probably the second branch of Bilateria to split off after the Acoelomorpha. Compared to its sister group, the omnipresent and spectacularly diverse Protostomia (containing the arthropods, molluscs and nematodes as its particular success stories), the deuterostomes are merely minor players. Interestingly, while protostomy has been generally regarded as ancestral to deuterostomy, modern phylogenies suggest the opposite, as shown by this phylogeny based on Giribet et al. (2000) where D = deuterostomous and P = protostomous:
While deuterostomy probably represents a secondary reversal in Brachiozoa other phylogenies place Brachiozoa nested within other Trochozoa, reinforcing this probability), protosotomous taxa as a whole are unmistakably nested within deuterostomous taxa.
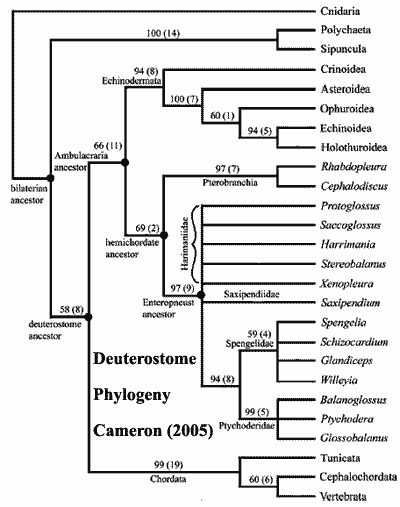
Within the Deuterostomia, molecular data have argued convincingly for a rearrangement of the main relationships (Cameron et al. 2000). The primary division of living members appears to be between the chordates on one hand and the echinoderms + hemichordates (forming a clade called Ambulacraria) on the other. This suggests that the characters shared between chordates and hemichordates may be ancestral for deuterostomes as a whole, and subsequently lost in echinoderms - a not-too-surprising state of affairs considering the greatly altered morphology of echinoderms. Heinzeller & Welsch (1999) regarded the hydrocoel of echinoderms as homologous with the notochord of chordates, as suggested by similarities in development and gene expression. The tornaria-type larva of Ambulacraria can no longer be assumed to be ancestral for Deuterostomia - it may represent an autapomorphy of Ambulacraria.
Recently, a molecular study has suggested that the obscure worm Xenoturbella might also fall within the Deuterostomia, specifically as the sister taxon to Ambulacraria. We therefore give it a look here. Also covered are the Vetulicolia, a group of Cambrian fossils that may represent the stem-group of crown deuterostomes, as well as a number of fossil groups regarded as stem echinoderms.
Not covered for now are a few other fossil groups that have been less certainly referred to Deuterostomia at one time or another, such as Odontogriphidae, Cambroclavida and Dinomischus. Also absent are the Chaetognatha arrow-worms), which have been considered the basalmost branch of the Deuterostomia in the past, but are currently regarded by many as basal protostomes (Helfenbein et al., 2004).
© Christopher Taylor, CT050815
Image credits: Deuterostomes picture not even remotely to scale. Komodo Lizard from Manfreds Travel Pictures. Dolichoglossus from the Bio 3650L Review Page (Clayton State University). Asterias amurensis from Kenya Yamamoto (Gifu University). Cameron (2005) cladogram from Cameron 2005).
One feature of the deuterostomes deserves a closer look - the widespread occurrence across the clade of deviations from the bilateral symmetry found in most Bilateria. Almost all the deuterostomes are asymmetrical to some degree, and in echinoderms bilateral symmetry has been lost entirely in the adult and replaced by pentaradial symmetry.
 In vertebrates, left-right asymmetry is mostly found in the viscera- we're probably all aware that our heart and digestive organs are not positioned directly down the midline, but are coiled and directed to one side or another depending on the organ. Other features, such as the skeleton, are more or less symmetrical.
In vertebrates, left-right asymmetry is mostly found in the viscera- we're probably all aware that our heart and digestive organs are not positioned directly down the midline, but are coiled and directed to one side or another depending on the organ. Other features, such as the skeleton, are more or less symmetrical.
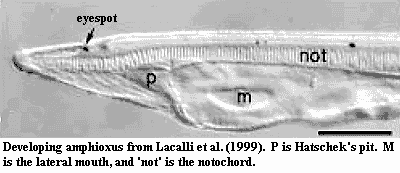
The immediate outgroup of craniates, the Cephalochordata e.g., amphioxus), show strong asymmetry during development. From the appearance of the fifth somite, the anterior borders of the somites move visibly out of register. Minguillon & Garcia-Fernandez (2002); Boorman & Shimeld (2002). After neurulation, paired cavities called Hatschek's diverticula (possible pituitary homologue) form in the head endoderm. The left cavity moves posteriorly to form the pre-oral pit, while the right expands to fill the vacated space in the head. The larval head becomes significantly asymmetrical, with the mouth appearing on the left side and the gill slits (originating on the ventral midline) spreading up to right side only. At metamorphosis, the mouth moves to the front, the pre-existing  gill slits move from the right side to the left, and new gill slits open on the right. The adult therefore regains pharyngeal symmetry. Other asymmetries remain, such as the displaced muscle blocks, the position of Hatschek's pit (an organ derived from the pre-oral pit, and probably homologous to the pituitary gland) on the left side, and a blind gut diverticulum on the right (Boorman & Shimeld, 2002).
gill slits move from the right side to the left, and new gill slits open on the right. The adult therefore regains pharyngeal symmetry. Other asymmetries remain, such as the displaced muscle blocks, the position of Hatschek's pit (an organ derived from the pre-oral pit, and probably homologous to the pituitary gland) on the left side, and a blind gut diverticulum on the right (Boorman & Shimeld, 2002).
Urochordata, the next group out, show little asymmetry in the larva, except in the positions of the two sensory pigment spots and adjoining regions of the brain. In the adult, the gut becomes asymmetrically folded.
The hemichordates are the most symmetrical of the deuterostomes. The gut is coiled asymmetrically but unlike other deuterostomes the direction of asymmetry differs between individuals. Directional asymmetries are few and relatively small - the protocoel pore is typically on the left. Boorman & Shimeld 2002). Rhabdopleura is something of an exception - in this genus, the right feeding arm is usually longer than the left, the single gonad is on the right, the mouth is on the left and the anus is on the right. Jefferies 1986).
In echinoderms the larva is initially bilaterally symmetrical, only to be replaced by a whole new order of symmetry in the adult. In sea urchins the pentaradial adult body plan is laid down in a structure called the adult rudiment, which lies on the left within the larva. Boorman & Shimeld (2002); Arenas-Mena et al. (2000). In starfish and crinoids, the left somatocoel becomes oral and the right one aboral. In starfish, only the left hydrocoel of the larva develops into the adult water-vascular system, while the right hydrocoel disappears. Jefferies (1986). A lot of debate still occurs about the appropriate interpretation of echinoderm axes, and how (if at all) they can be compared to axes in other Bilateria. It is uncertain, for instance, if the anterior-posterior axis in other animals corresponds to the oral-aboral axis of 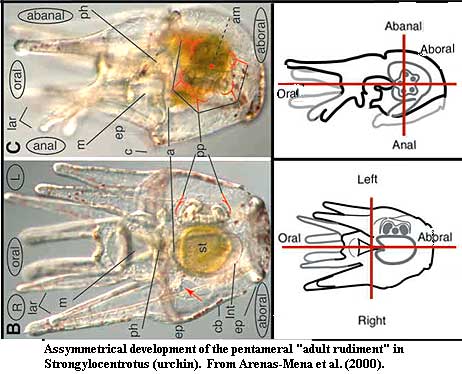 echinoderms or the central axes of the five rays. The fossil lineage of echinoderms suggests some very odd evolutionary stages, with homalozoans representing a strongly asymmetrical stage in echinoderm ancestry (if they are indeed stem echinoderms - see later).
echinoderms or the central axes of the five rays. The fossil lineage of echinoderms suggests some very odd evolutionary stages, with homalozoans representing a strongly asymmetrical stage in echinoderm ancestry (if they are indeed stem echinoderms - see later).
The obvious question then becomes what this all means. Why are deuterostomes such individualists? Jefferies (e.g., 1986) has suggested that the asymmetry of chordates and echinoderms is homologous, and evidence that they form a monophylum to the exclusion of the more symmetrical hemichordates. This clade he named Dexiothetica after his suggested explanation for its origin - that the ancestor of the echinoderm-chordate clade underwent a 90 degree rotation so that it came to lie on its right side with the left side becoming dorsal (Greek dexios = right, thetikos = suitable for lying down). In chordates, the outer body subsequently re-oriented itself over time to become bilaterally symmetrical, but the viscera remained dexiothetic.
Jefferies' theory was strongly dependent on his interpretation of the fossil homalozoans (which he referred to as calcichordates - see later). On this page, we take the more orthodox view that homalozoans are stem echinoderms, as suggested by their possession of a calcite skeleton with a similar ultrastructure to modern echinoderms. Jefferies held this skeleton to be ancestral for Dexiothetica, and subsequently independently lost in each of the three chordate subphyla. However, this seems to require a lot of convergence, expecially if, as molecular phylogenies suggest, the hemichordates are actually closer to echinoderms. A stem echinoderm position for homalozoans seems much more parsimonious.
Could dexiothetism have still been ancestral to deuterostomes as a whole? The theory does have a certain appeal in relation to echinoderms - as mentioned before, left and right do become dorsal and ventral in echinoderm development, and the unpaired nature of the gill structures in Vetulocystidae and Cornuta at the base of the echinoderm stem offers further support (if they are correctly interpreted as such - see later). In the case of Chordata, however, if the homalozoans are not regarded as relevant to the chordate stem, the support for dexiothetism is much lower. Asymmetries in the viscera may have been an adaptation for greater complexity (a coiled heart has greater pumping ability, while a coiled gut allows for more surface area in a smaller volume. Cooke 2004). The strong asymmetry in amphioxus may be an autapomorphy of that particular lineage. Evidence is therefore equivocal as to whether dexiothetism can be regarded as ancestral for Deuterostomia, or whether chordate and echinoderm asymmetries arose independently. Among fossils that may lie along the stem group of deuterostomes, the Vetulicolia have not been recorded as showing any sign of asymmetry. However, the mostly lateral, two-dimensional preservation of vetulicolians means that this is not necessarily informative.
Studies of developmental gene expressions and their relation to deuterostome asymmetry are still at a fairly early stage. The Nodal-Ptx pathway has been identified as a major factor in establishing the left-right axis in chordates. Ptx, for instance, is expressed predominantly on the left-hand side of the embryo in all three chordate subphyla. However, neither Pitx or Nodal have yet been identified in echinoderms, though once again the extremely derived body plan of echinoderms makes the identification of gene functional homologies difficult. Wray & Lowe (2000). Preliminary analysis suggests Ptx expression is symmetrical in hemichordates, but this requires confirmation. Boorman & Shimeld (2002).
In succeeding pages, we will explore some of the strange fruits that have blossomed on the Deuterostome stem. For now, we include some other, quite different, versions of Deuterostome phylogent for comparison.
© Christopher Taylor, CT050821.