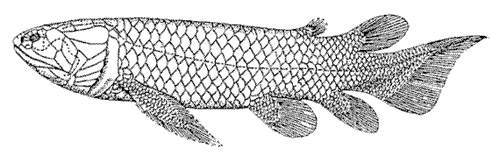
Holoptychius - late Devonian period
| Sarcopterygii | ||
| The Vertebrates | Overview |
| Vertebrates Home | Vertebrate | Vertebrate |
|
Abbreviated Dendrogram
Teleostomi
├─Neopterygii
└─Sarcopterygii
├─┬─Onychodontiformes
│ └─Actinistia
└─Rhipidistia
├─Dipnomorpha
└─┬─Rhizodontiformes
└─Osteolepiformes
├─Tristichopteridae
└─Elpistostegalia
├─Panderichthys
└─Tetrapoda
|
Contents
Overview |
|
|
Holoptychius - late Devonian period |
The Sarcopterygii. are generally robust osteichthyan fishes with fleshy lobe-fins. They are distinguished from the Actinopterygians or ray-finned fish by their muscular fins with articulating bones. The fins are called lobe fins because they are fleshy and like the limbs of land animals, rather than the fins of familiar "ray-finned" fish. The lobe fins swivel on a shoulder or hip socket (like a tetrapod limb) and most of the bones can be matched directly to the bones in the limbs of land animals. Even the individual rays in the fin can be moved individually, like toes. It is easy to see how everything fit together in giving the Sarcopterygii a transitional role between jawed fish and the Tetrapoda.
There are traditionally three main groups of Sarcopterygii, the Crossopterygia no longer regarded as a valid taxon), the Coelocanths, once an important group but now limited to a single large deepwater genus, and the lungfish (Dipnoi), which are capable of breathing air and surviving out of water. In fact all sarcopterygians have lungs of some sort, as do even primitive Actinopterygia and some Placoderms. Air-breathing in fish is, in fact, a primitive character of all osteichthyans.
Sarcopterygii are also distinguished by a dermal or external (scaly) skeleton characterized by the shiny dermal tissue cosmine, a feature secondarily lost in all advanced lineages, and by the presence of unique tooth materials, such as enameloid on the teeth of predatory forms, and mineralized dentines, such as petrodentine, in the toothplate and denticles of lungfishes.
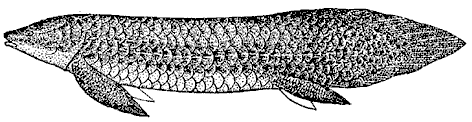 The Sarcopterygii first appeared at least as early as the latest Silurian period. The earliest well-known forms are from the Early Devonian and include forms like Youngolepis, and Powichthys, whose affinities are completely unclear. They are variously considered ancestral or "stem-group" sarcopterygians from which the other forms evolved, or primitive proto-lungfish. From the ancestral types the sarcopterygians quickly diversified into a diverse group of mostly large fish as the Devonian period progressed. They flourished in marine, brackish and fresh water environments and made up an important part of the Devonian fauna. They were hit hard by the terminal Devonian extinction event, and most marine forms died out then, only the coelacanths continuing. Freshwater types continued to flourish, including the rhizodonts and lungfish. The rhizodonts were large predators, sometimes attaining a length of several meters. By the Mesozoic, the coelacanths were less common in the seas, and only the lungfish remained as important inhabitants of rivers and lakes. Living lungfish are virtually unchanged from their Mesozoic ancestors. The living Australian Neoceratodus left) in particular is a true "living fossil". Today only a few species of extant lungfish (Dipnoi) and the coelacanth Latimeria remain, the last survivors of a once flourishing group.
The Sarcopterygii first appeared at least as early as the latest Silurian period. The earliest well-known forms are from the Early Devonian and include forms like Youngolepis, and Powichthys, whose affinities are completely unclear. They are variously considered ancestral or "stem-group" sarcopterygians from which the other forms evolved, or primitive proto-lungfish. From the ancestral types the sarcopterygians quickly diversified into a diverse group of mostly large fish as the Devonian period progressed. They flourished in marine, brackish and fresh water environments and made up an important part of the Devonian fauna. They were hit hard by the terminal Devonian extinction event, and most marine forms died out then, only the coelacanths continuing. Freshwater types continued to flourish, including the rhizodonts and lungfish. The rhizodonts were large predators, sometimes attaining a length of several meters. By the Mesozoic, the coelacanths were less common in the seas, and only the lungfish remained as important inhabitants of rivers and lakes. Living lungfish are virtually unchanged from their Mesozoic ancestors. The living Australian Neoceratodus left) in particular is a true "living fossil". Today only a few species of extant lungfish (Dipnoi) and the coelacanth Latimeria remain, the last survivors of a once flourishing group.
Here is a double cladogram of Sarcopterygian interrelationships, from Janvier (1996). The right and left side summarize two rival interpretations (and there are others). The left-hand side is one of the fully resolved cladograms, essentially from Ahlberg (1991a), which we follow here.
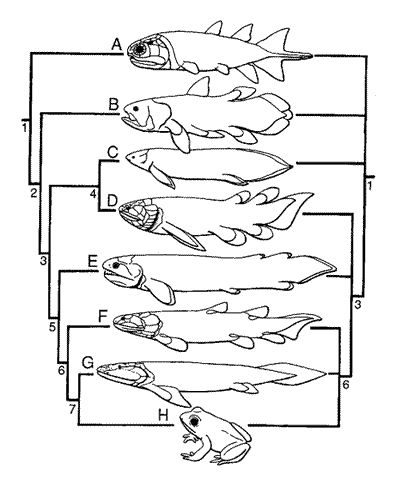 |
Terminal taxa:A, Onychodontiformes - early to late DevonianB, Actinistia - middle Devonian to Recent C, Dipnoi - late Silurian or early Devonian to Recent D, Porolepiformes - early to late Devonian E Rhizodontiformes - late Devonian to Carboniferous F. Osteolepiformes - middle Devonian to early Permian G. Panderichthys - ?middle to late Devonian H. Tetrapoda - late Devonian to Recent Nested taxa and selected synapomorphies (unique characteristics):1 Sarcopterygii (monobasal paired fins, ventral fissure developed dorsally into an intracranial joint, more than four scelerotic plates); 2, maxilla reluced or lost, large basal lobe in fins; 3, Rhipidistia folded dentine and enamel in teeth, no maxilla-preopercular contact, large quadratojugal. postorbital junction of supraorbital and infraorbital sensory-lines [lost in lungfishes], only three coronoids, nasal capsule with lateral recess, ?dermintermedisl process [lost in porolepiforms]); 4, Dipnomorpha (elongated pectoral fin, with numerous mesomeres, rostral tubules, branched posterior radials in posterior dorsal fins, enamel entering the flask-shaped cavities of cosmine, pineal reached but not surrounded by parietals, no parietal-supraorbital contact, infraorbital sensory-line canal follows premaxillary suture); 5, Tetrapoda (concave glenoid fossa, perforated hurneral process, large entepicondylar process, deltoid and supinator processes on humerus); 6, Choanata (single external nostril, choanae. dermintermedial and tectal processes in external nostril [unless at node 3]); 7. large frontals meeting along the midline, elongated humerus, jugal meeting quadratojugal, dorsally placed orbits, ventrally placed external nostril near oral margin, loss of dorsal and anal fins) |
MAK 000114.
checked ATW051001