Ornithischia: Hadrosauroidea
Taxa on This Page
- Altirhinus X
- Hadrosauroidea X
- Nanyangosaurus X
- Ouranosaurus X
Mesozoic Cattle or Mesozoic Camels?
 Iguanodon may be one of those rare cases in which we actually see the base of a significant radiation. Near - iguanodonts like Ouranosaurus and Nanyangosaurus may or may not turn out to be hadrosauroids, i.e. slightly more closely related to hadrosaurs than to Iguanodon. They are certainly very closely related to the iguanodontids; and the differences, if any, are tenuous. For example, when data from Head (2001) is run on PHYLIP, there are no synapomorphies which distinguish hadrosauroids from iguanodontids. Ouranosaurus comes out as the most basal hadrosauroid, but it is at least as different from the other basal hadrosauroids as it is from iguanodontids. [1] At a guess, Ouranosaurus will turn out to be a rather derived member of an early, African offshoot from the iguanodontid stem. However, despite much new data and a great deal of careful work over the last twelve years, notably by David Norman, these dinosaurs are all so similar to each other that we are not a great deal closer to understanding exactly where the branching occurs than when The Dinosauria was published in 1990. See, Weishampel & Horner (1990); Norman (1990). One significant problem may be the number of species referred to Iguanodon. The genus is overdue for revision. [2] But who has time and resources for so massive an undertaking?
Iguanodon may be one of those rare cases in which we actually see the base of a significant radiation. Near - iguanodonts like Ouranosaurus and Nanyangosaurus may or may not turn out to be hadrosauroids, i.e. slightly more closely related to hadrosaurs than to Iguanodon. They are certainly very closely related to the iguanodontids; and the differences, if any, are tenuous. For example, when data from Head (2001) is run on PHYLIP, there are no synapomorphies which distinguish hadrosauroids from iguanodontids. Ouranosaurus comes out as the most basal hadrosauroid, but it is at least as different from the other basal hadrosauroids as it is from iguanodontids. [1] At a guess, Ouranosaurus will turn out to be a rather derived member of an early, African offshoot from the iguanodontid stem. However, despite much new data and a great deal of careful work over the last twelve years, notably by David Norman, these dinosaurs are all so similar to each other that we are not a great deal closer to understanding exactly where the branching occurs than when The Dinosauria was published in 1990. See, Weishampel & Horner (1990); Norman (1990). One significant problem may be the number of species referred to Iguanodon. The genus is overdue for revision. [2] But who has time and resources for so massive an undertaking?
(Perhaps some misguided reader is, even now, racing out his door in an attempt to become the answer to this entirely rhetorical question. The probability of such an occurrence is, admittedly, on the same order as the probability of spontaneous combustion. While intriguing to contemplate -- everyone knows of at least one paleontologist who is worthy of immolation (spontaneous or otherwise) -- such an eventuality need not detain us long. The chance that your editors can contribute anything meaningful to the discussion of basal hadrosauroid phylogeny is thus remote. Like a hypothetical bull Parasaurolophus in rut, nature has equipped us to make impressive and sometimes attractive noises, but enjoined us to leave the field of actual combat to those better equipped for offensive action.)
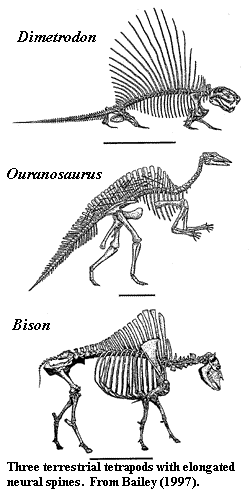
Although its ultimate antecedents are somewhat vague, Ouranosaurus remains a fascinating subject. Phillipe Taquet, as he relates in an autobiography, spent much of his early career on the discovery and reconstruction of this dinosaur from the middle Cretaceous of Niger. Taquet 1998). [3] Not only does Ouranosaurus come from an unusual locale, but some of its anatomy is downright weird. Most notably, Ouranosaurus has very tall neural spines on its dorsal vertebrae. These elevated spines are tallest above the forelimbs, but extend all the way down to the first few caudals. Tall neural spines are not particularly unusual among dinosaurs, but the spines of Ouranosaurus are much longer than anything else in the phylogenetic neighborhood. For that reason, Ouranosaurus has customarily been illustrated as having a sailback, with the sail having some presumably thermoregulatory function, like Dimetrodon. See Ouranosaurus- Enchanted Learning Software; OURANOSAURUS.
This enduring image of the sailback hadrosauroid is difficult to explain. Bailey 1997) makes an airtight case that the spines have no resemblance to the relatively few terrestrial sailbacks known from tetrapod zoology and paleontology. texas dinosaurs; New Scientist | Dinosaurs. Rather, the spines are, in shape, size, distribution, mass and contour much closer to the hump-supporting spines of ungulates, most notably buffalo. [4] Like buffalo, the neural spines of many, perhaps all, iguanodontians were bound together by extensive tendons which stiffened the back and supported the head. We can be quite certain of this, since some of the tendons frequently ossified. Ouranosaurus is no exception. Consequently, the dorsal structure, whatever it was, could not have been fin-like. It had to be at least moderately thick transversely in order to enclose the tendons.
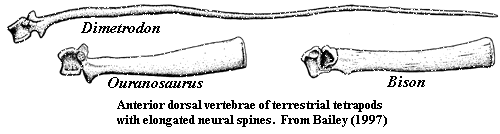 In bona fide sailbacks, the spine's only substantial function is to support itself and a vascular network. If the function of the dorsal structure is thermoregulatory, its efficiency is much greater with a maximum surface to volume ratio. The greater the surface, the more efficient is the heat exchange. Transverse width decreases efficiency. Accordingly, where the function of the sail is thermoregulatory, the spines in are not only thin, but can and do become thinner distally. The distal parts simply have less weight to support. This is the pattern seen in the dorsal fins of fishes, in Dimetrodon and among iguanid lizards. By contrast, Bison and Ouranosaurus have massive spines that actually become thicker distally. In such humped animals, the dorsal structure is wide, heavy and rather far off the ground. Unless the hump is tightly constrained, it can generate lateral torque which will cause the animal to tip over. Like an overloaded bus going around a mountain curve, a sudden shift in weight can topple the whole structure with disastrous results. Accordingly, both natural selection and prudent bus drivers tightly lash top-loaded baggage. Hadrosauroids did not come equipped with baggage racks. Instead, like subway cars, they had a series of central poles onto which everything held tightly when rounding a corner or accelerating quickly. Without question, hadrosauroids were quieter and more sanitary than subway cars, but the principle is the same.
In bona fide sailbacks, the spine's only substantial function is to support itself and a vascular network. If the function of the dorsal structure is thermoregulatory, its efficiency is much greater with a maximum surface to volume ratio. The greater the surface, the more efficient is the heat exchange. Transverse width decreases efficiency. Accordingly, where the function of the sail is thermoregulatory, the spines in are not only thin, but can and do become thinner distally. The distal parts simply have less weight to support. This is the pattern seen in the dorsal fins of fishes, in Dimetrodon and among iguanid lizards. By contrast, Bison and Ouranosaurus have massive spines that actually become thicker distally. In such humped animals, the dorsal structure is wide, heavy and rather far off the ground. Unless the hump is tightly constrained, it can generate lateral torque which will cause the animal to tip over. Like an overloaded bus going around a mountain curve, a sudden shift in weight can topple the whole structure with disastrous results. Accordingly, both natural selection and prudent bus drivers tightly lash top-loaded baggage. Hadrosauroids did not come equipped with baggage racks. Instead, like subway cars, they had a series of central poles onto which everything held tightly when rounding a corner or accelerating quickly. Without question, hadrosauroids were quieter and more sanitary than subway cars, but the principle is the same.
Bailey compares the environment and ecology of Ouranosaurus to the environment and ecology of ungulates, notably camels. He argues that in the Cretaceous, as today, Niger was at least seasonally arid, semi-desert, open country with a low browse line. In addition to fat storage for water reserves, Bailey speculates that the humps may have provided massive food reserves required for migration to isolated nesting or nursery areas, as well as providing tendon attachments for the forelimb muscles to allow more economical long-distance walking.
Although this sort of analogy to herd-living, open-country ungulates will always be somewhat conjectural, it is far from being completely speculative. Considerable evidence supports the idea (which may originally have been advanced by Peter Dodson) that hadrosauroids were the "ungulates of the Mesozoic." A recent study by Carrano et al.. (1999) attempted to test the hypothesis in a quantitative manner, using multivariate analysis of morphometric data. The results were not conclusive, but were encouraging. It is not beyond hope that this generalized hypothesis about hadrosauroid lifestyles can be tested. ATW020215
[1] Under these conditions, the apomorphies of Ouranosaurus are (a) lateral expansion of the rostrum; (b) dorsoventral expansion of the anterior jugal; (c) large mandibular diastema (d) ventral ridge on sacral vertebrae; and (e) an anteroposteriorly narrow supratemporal fenestra. All of these characters are convergent on various hadrosaurids. Back.
[2] See the discussion in the notes following Hadrosauroidea. Back.
[3] Taquet's monograph was published as: Taquet, P 1976), Geologie et paleontologie du gisement de Gadoufaoua (Aptien du Niger). Cah. Paleontol. CNRS Paris. 191 pp. However, I have not reviewed the original work. Back.
[4] Note that the Bison shown in the image is not the living B. bison, but its somewhat larger Pleistocene relatives, B. antiquus (skeletal reconstruction) and B. latifrons (isolated vertebra). Back.
Descriptions
Hadrosauroidea:
Probactrosaurus, Altirhinus. Definition: Hadrosaurus > Iguanodon.
Phylogeny: Iguanodontia :::: Iguanodontidae + * : Ouranosaurus + (Nanyangosaurus + (Altirhinus + Hadrosauridae)).
Characters: $? laterally expanded distal rostrum ("duck bill") [H98] [N90* (synapomorphy of Iguanodontia)]; $ narial fossa lengthened and defined laterally by raised rim [N90* (present in Iguanodon and absent in lambeosaurines)]; $ postorbital skull narrow [N90* (also in iguanodontids)]; $ squamosals approach midline of skull roof, separated only by narrow band of parietal [N90* (same in Iguanodon)]; $ anterior end of jugal dorsoventrally expanded under orbit [N90*]; distal end of paroccipital process curves anteriorly [N90*]; $ enlarged diastema [N90*]; tooth size reduced compared to iguanodontids [H98]; $ 1+ maxillary and dentary tooth per tooth position [N90]; most have 3+ dentary teeth per tooth position [H98]; most have maxillary teeth symmetrical around a medial carina (but see [1]) [H98]; maxillary teeth without mesial & distal accessory ridges [H98]; $ parallel, symmetrical accessory ridges on dentary teeth [N90]; $ caudal neural spines longer then chevrons [N90*]; $ scapular blade with convex dorsal margin [N90* (not true of Ouranosaurus)]; $ ilium with small pubic peduncle [N90* (synapomorphy of higher-level clade within Iguanodontia)]; $ ilium with antitrochanter [N90* (denies that Ouranosaurus antitrochanter is homologous)]; $ ilium with long preacetabular process [N90* (same in one species of Iguanodon)]; $ ischium straight [N90]; $ dorsoventral expansion of prepubic process [N90* (synapomorphy of higher-level clade within Iguanodontia)]; $? loss of metatarsal I [N90] $ distal tarsals 2&3 absent [N90* (may be fused or not)]; pes digit I absent [N90*].
References: Head (1998) [H98]; Norman (1990) [N90].
Notes: [1] i.e., the tooth has a ridge running from base to crown and the teeth are symmetrical around that axis in lingual view. Head's (1998) description is confusing on this point. The text states that the maxillary teeth have a "single medial" carina. His Figure 13(a), however, purports to be in buccal view and shows a prominent median carina -- a carina which is in the middle, but on the lateral or buccal (not medial or lingual) face. Is the figure mislabeled or is medial a typographical error? See discussion at median. [2] Norman (1990) places Probactrosaurus as the sister of Hadrosauridae, and Ouranosaurus as an iguanodontid. Accordingly, the characters from [N90] actual characterize a different, unnamed group: Probactrosaurus + Hadrosauridae. [3] Characters marked [N90*] are from Norman's discussion of prior work by Sereno. See Sereno, PC (1986), Phylogeny of the bird-hipped dinosaurs. Natl. Geog. Res. 2: 234-256. Norman disagrees, and gives detailed, well thought-out reasons, some of which are noted above. The argument may turn on the proper interpretation of I. atherfieldensis, which Norman repeatedly uses as a counter-example to the synapomorphies he cites to Sereno. It is tempting to speculate that atherfieldensis might not be a species of Iguanodon after all. Norman himself has shown that I. orientalis (now Altirhinus) is actually a hadrosauroid, and perhaps the same is true of I. atherfieldensis. Alternatively, perhaps Iguanodon is paraphyletic. ATW020202.
 Ouranosaurus:
Ouranosaurus:
Range: Middle Cretaceous of Africa
Phylogeny: Hadrosauroidea : Nanyangosaurus + (Altirhinus + Hadrosauridae)) + *. [H01] [2]
Characters: large bodied (7 m); low skull; broad, flat rostrum (duck bill) [H98] [N90]; lateral margins of nares widely flared [NW90]; long, sloping facial region [NW90]; premaxilla lateral processes reconfigured to support nasals [NW90]; anterior maxilla reconfigured as shelf to support premaxilla (as in lambeosaurines, but likely convergent) [H98] [NW90]; marked free palpebral; crest of "bumps" on nasals near frontal suture [NW90]; prefrontal more dorsal than in iguanodontids [NW90]; marked free palpebral; long axis of supratemporal fenestra oval & oriented anterolaterally [WX01]; anterior jugal somewhat dorsoventrally expanded (convergent with Hadrosauridae?) [H98]; quadrate shaft curved [WX01]; supraoccipital may be excluded from foramen magnum [NW90]; occipital condyle almost heart-shaped (seems inconsistent with figure) [NW90]; braincase low & broad as in hadrosaurids [NW90]; predentary very broad [NW90]; dentary with straight dorsal & ventral margins [WX01]; prominent diastema between predentary and 1st dentary teeth [H98]; <3 dentary teeth per tooth position [H98]; maxillary with 22 tooth positions, dentary with
23 [NW90]; maxillary tooth crowns with accessory mesial & distal ridges [H98]; tooth ultrastructure showing numerous odontoblasts and large canals parallel to odontoblast tracks (similar to certain living ungulates) [M88] [1]; vertebral formula 11/17?/6/40+ [NW90]; neck short; long (9x height of centra [NW90]) neural spines possibly supporting "sail" or, more likely, hump [B97]; spines robust, eliptical with long axis parallel to body axis [B97]; spines variably bowed, slanted and/or with expanded distal ends (all correlated with strong, distally applied, anteroposterior strains as in Bison) [B97]; neural spines interconnected by (partially?) ossified tendons [B97]; hoof-bearing digits 2&3 on forelimbs; manus 1 with spike [NW90]; ischium shaft relatively straight [N90]; pes with phalangeal formula 03450 [NW90]; low browser in open, arid or seasonally arid environments [B97] .
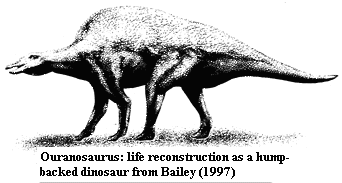 Links: DinoData: Ouranosaurus; Ouranosaurus- Enchanted Learning Software; Ouranosaurus; Ouranosaurus also in French); Ouranosaurus The Natural History Museum's Dino Directory; Ouranosaurus Finished; FPDM : Ouranosaurus nigeriensis; Ouranosaurus; Dinosaur Impressions by Taquet; dinos.htm;
Links: DinoData: Ouranosaurus; Ouranosaurus- Enchanted Learning Software; Ouranosaurus; Ouranosaurus also in French); Ouranosaurus The Natural History Museum's Dino Directory; Ouranosaurus Finished; FPDM : Ouranosaurus nigeriensis; Ouranosaurus; Dinosaur Impressions by Taquet; dinos.htm;
References: Bailey 1997) [B97]; Head (1998) [H98]; Head 2001) [H01]; Michard (1988) [M88]; Norman (1990) [N90]; Norman & Weishampel (1990) [NW90]; Wang & Xu (2001) [WX01].
Notes: [1] Michard (1988) describes these tubules as having an inner covering including odd, circular structures. For a variety of reasons, I suspect these are artifacts. However, it is interesting that the "tubules géants" were not filled with matrix as were the odontoblast tubules -- or were filled with something which was destroyed by the acid) preparation for scanning electron microscopy, leaving only the circular structures. Michard does not believe these are vascular canals (blood vessels) because of their irregular distribution and because they are surrounded by concentric layers of dentine. However, they may be the remains of a temporary vessel network, most of which was resorbed during tooth development. This would be consistent with the very long odontoblast tracks. How else could the odontoblasts be fed and their wastes removed during their long migration? Michard observes that the density of these tubules (where present) was much greater near the pulp cavity -- again consistent with a vascular role. Michard states that comparative work was under way by various other workers, but I have not seen it. [2] Head's (2001) cladogram is ambiguous. When I run his data set, Ouranosaurus comes out as a hadrosauroid, but, as Head notes, the synapomorphies in this part of phylospace are weak and rather unconvincing. [3] See also discussion at Mesozoic Cattle? ATW020208.
Nanyangosaurus:
Range: Lower? or Middle Cretaceous of China.
Phylogeny: Hadrosauroidea :: (Altirhinus + Hadrosauridae) + *.
Characters: length about 4.5 m; 6 sacral vertebrae; caudal neural spines larger than chevrons; front and hind limbs similar to hadrosaurids; at least one anterior dorsal vertebra slightly opisthocoelous; all posterior dorsals platycoelous; dorsal centra with heart-shaped ends; some dorsals fused, lacking ventral keel (pathological?); 6 "true" sacrals, with one additional vertebra at each end associated with sacrum; sacral centra flattened & wide; caudal vertebrae platycoelous; ventral groove on posterior sacrals; caudal vertebrae progressively lose diameter faster than length, so most distal caudals are elongate; forelimb slender; distal humerus dorsoventrally expanded; metacarpal I absent; metacarpals II, III & IV partially fused; metacarpals III & IV robust with expanded distal end on IV; phalanges robust; manual phalanx IV(1) wider than long; unguals hoof-like; distal ischium moderately curved with well-developed foot; femur straight in lateral view; femur with medial distal condyle more than twice as wide as lateral; anterior intercondylar groove is tunnel-like; distal femoral condyles not fused; proximal tibia transversely wide; fibula semilunate in proximal view; astragalus & calcaneum fused and rectangular anteriorly with a weak ascending process; metatarsal I absent, metatarsal II deeper proximally than metatarsal III; pedal unguals hoof-shaped.
Links: DinoData: Nanyangosaurus; Nanyangosaurus -- The Dinosauricon; Dinosauria Translation and Pronunciation Guide N; nanyangosaurus.
References:Norman 1990).
Notes: [1] the skull is not known. [2] According to Norman (1990), variable concavity of the centra and centra with heart-shaped ends are characters of immature specimens of Iguanodon. They both result from incomplete ossification -- of the centra and neural arch pedicles, respectively. Immature Iguanodon likewise exhibit sacrodorsals which become true sacrals in mature specimens. It seems unclear what, if anything, really distinguishes Nanyangosaurus from Iguanodon. ATW020201.
 Altirhinus: = Iguanodon orientalis.
Altirhinus: = Iguanodon orientalis.
Range: Middle Cretaceous of Mongolia
Phylogeny: Hadrosauroidea :: Hadrosauridae + *.
Characters: length up to ~8m [NS00]; dorsally expanded rostrum; beak wide and anteroventrally oriented [NS00]; nasal strongly flexed dorsally [NS00]; nasals with elongate rostral process [NS00]; internasal groove present [NS00] (could be cartilaginous extension of premaxilla?); antorbital fenestra "occluded" (covered by frontal in lateral view?) [1] [NS00]; dentary deflected ventrally [NS00]; posterior dentary and maxilla with 3 teeth in each vertical tooth row [NS00]; carpals not fused [NS00]; manus I with laterally compressed ungual spike [NS00](as Iguanodon?); manus IV with short, broad ungual [NS00]; ilium with thick, downturned anterior process [NS00]; brevis shelf absent [NS00]; ilium with strong trochanteric ridge above body of ischium [NS00] shaft of ischium straight, but with axial twist [NS00].
Links: DinoData: Altirhinus; Altirhinus; Altirhinus Fact Sheet - EnchantedLearning.com; Witmer's Lab Dinosaur Skull Collection: Other Ornithischia; Altirhinus Portuguese); Dinosauria Translation and Pronunciation Guide A.
Image: Altirhinus
References: Norman 1990), Norman & Sues (2000) [NS00]
Notes: [1] this section of the frontal is broken off in the image. [2] The description of the ilium differs only a little from the description of Iguanodon in Norman 1990). However, Iguanodon has a narrow brevis shelf, and the shaft is twisted outward. Id. 020201.
{kind=link}