
| Aves | ||
| The Vertebrates | Overview |
| Vertebrates Home | Vertebrate | Vertebrate |
|
Abbreviated Dendrogram
Coelurosauria
│
└─Aves
├─Archaeornithes
└─┬─Confuciusornithidae
└─Ornithothoraces
├─Enantiornithes
└─Ornithurae
├─Hesperornithiformes
└─Neornithes
├─Paleognathae
│ ├─Lithornithiformes
│ └─Ratites
└─┬─Galloanserae
└─Gruimorpha
|
Contents
Index |
The first protobirds (Archaeornithes) evolved from small Coelurosaurian theropods during the late Jurassic (probably Kimmeridgian age). They were feathered gliders or jumpers and weak flyers that may have taken to the trees in to avoid ground-based predators. The mid-Tithonian extinction event eliminated a lot of the big dinosaurs and produced some empty ecological niches. Some Archaeornithines exploited these niches by becoming secondarily flightless ground-based predators and omnivores. Others however evolved into more efficient and advanced fliers. Thus for the first five or ten million years of the Cretaceous there would have been a tremendous evolutionary radiation. The result was about half a dozen or more different orders of birds, ranging from very dinosaur-like to very (modern-) bird-like, coexisting and co-evolving together throughout the seventy or eighty million years of the Cretaceous.
The terrible extinction event at the end of the Mesozoic (possibly a large asteroid impact, preceded and succeed by an extended period of vulcanism with consequent nuclear winter, acid rain etc) put an end to this primordial garden of Eden. Many protozoa and invertebrates as well as dinosaurs, pterosaurs, marine reptiles, and a number of bird and mammal lineages were taken out by this traumatic extinction. The only lineage of endothermic archosaurs to survive into the Cenozoic were the Neornithes or "modern birds". These likewise underwent a huge evolutionary radiation during the early Tertiary, including some amazing secondarily flightless forms. They were however never as anatomically or probably) metabolically diverse as the Cretaceous protoaves and veraves. Today the Neornithes remain one of the largest and most successful groups of tetrapods. MAK991003.
| |
|
|
|
The transition from basal Dinosauria to Aves was relatively gradual. The missing links have become fairly small. In fact, the characters separating birds and other Maniraptora are astonishingly minor. However, the morphological distance between dinosaurs and neornithine birds is still impressive, and modern birds are extremely specialized. The following is a description of a generalized neornithine bird, emphasizing the terminology unique to ornithology as well as the unique specializations of the avian form as it eventually evolved.
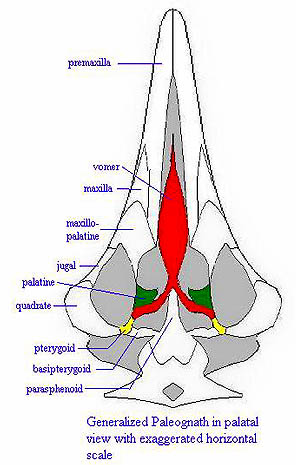
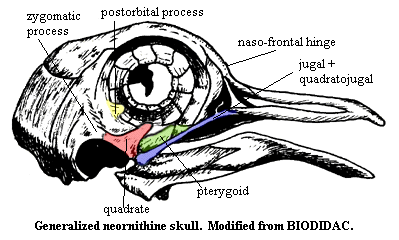
Skull: Loss of teeth; jaw function is taken over by keratinous rhamphotheca. The quadrate is highly mobile. Compared to more basal dinosaurs, birds have very large brains and orbits. The upper jaw is moveable on the braincase. Almost all of the dermal skull is fused. The upper beak is hinged dorsally on a naso-frontal hinge. Ventrolaterally, the upper beak has a long buttress (the jugal bar) to the quadrate, which articulates on a ventrolateral zygomatic process of the braincase. The quadrate, in turn, is braced by long, thin pterygoids which proceed anteromedially to meet on a fused medial bar that joins the palatine to the braincase. The palatines have anterior processes which meet and fuse with the upper beak anterior to nostrils. In the "vacuity" between these processes there are 2 small maxillo-palatines and 2 even smaller vomers. The lower jaw articulates directly on the quadrate, but is also connected by soft tissue to a post-orbital process of the braincase which lies dorsal to the zygomatic process.
Vertebral column: The cervicals typically bear small, posteriorly-directed ribs and are not interlocked or stiffened. The dorsals may be centrally fused to form a notarium. The dorsal ribs are double-headed and bear uncinate processes which overlap the next posterior rib. The dorsal ribs articulate with ends of sternal ribs which attach to the sternum. The sternum may incorporate additional vertebrae and is referred to as a synsacrum. The tail is shortened and fused to form a pygostyle (see BIODIDAC) which bears tail feathers (retrices). image
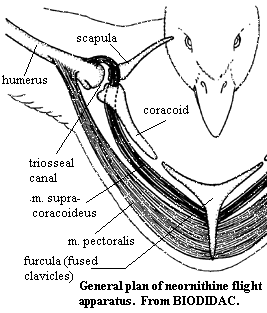
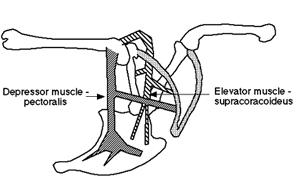
Anterior Limb girdle The anterior girdle is lateral and raised almost to level of the dorsal vertebrae. The scapulae are thin and extend almost directly posterior, almost covering the heads of the notarial ribs in lateral view. The coracoids extend ventro-medially and articulate broadly with the large sternum. The sternum runs postero-ventrally and bears a large keel (carina) which extends ventrally or antero-ventrally. The sternum also bears a posterolateral process which articulates with the sternal ribs. The outer curve of the carina more or less follows or defines the outer wall of the body. The scapulae are curved antero-medially from the dorsal end of the coracoids and meet and fuse at roughly the anterior-most portion of the breast. They may extend further ventrally to form an enlarged, flattened hypocleidium. The supracoracoideus muscle attaches more or less to the basal internal or ventral) sides of the carina. The scapulocoracoideus tendon passes dorsally between the coracoid and clavicle, and through the triosseal canal, to attach on the dorsal surface of the humerus and effect the recovery stroke in flight. The pectoralis muscle attaches on the more distal portions of the sternum and on the hypocleidium and inserts on the ventral (or anteroventral) portion of the humerus to power the flight stroke.
Wings: (see below & right, from Aves031B&File_type=GIF">BIODIDAC) The humerus is directed postero-laterally; the radius & ulna are directed antero-laterally and are bowed slightly away from each other. The ulna is posterior and bears the secondary flight feathers, numbered from 1 counting proximally beginning at the wrist). The radius and ulna terminate on small radiale and ulnare, respectively. The carpometacarpus is directed postero-laterally with a phalangeal formula 1+2+1. Digit 2 is large. The carpometacarpus and digits 2 & 3 bear the flight primaries, numbered from I (counting distally from proximal base of carpometacarpus). Digit 1 consists of a small alula which emerges from the proximal base of the carpometacarpus. The vinculum tendon emerges from the distal end of the humerus and follows the posterior margin of the wing, attaching at the distal end of digit 2. The raches of the remiges (i.e., the primaries and secondaries) are stabilized by passing through the vinculum. Another major tendon emerges from the proximal base of the humerus and inserts on the proximal base of the carpometacarpus. It thus forms the base of a triangle, the other 2 sides of which are formed by humerus and radius. This area is filled by a flap of skin (the prepatagium) and by muscle. One or more additional layers of shorter feathers may overlie the proximal parts of the flight primaries and secondaries. These are referred to as coverts.
Legs: (see figure at right from Aves029B&File_type=GIF">BIODIDAC; see also Aves028B&File_type=GIF">BIODIDAC) The pelvic girdle is relatively unspecialized. The pelvic bones are frequently fused in various ways, including distal posterior fusion of the ischium, ilium and/or pubis. However, the overall structure is not novel. The tibia and fibula are fused, especially distally, to form a single element, the tibiotarsus. Similarly, the tarsus is normally a single element, the tarsometatarsus. The structure of the foot is variable and often diagnostic. The basic avian foot is tetradactyl. The most general form is anisodactyl, with digits 2-4 anterior and 1(the hallux) reversed as a perching adaptation (see Aves010B&File_type=GIF">BIODIDAC).. Either 2 (heterodactyl) or 4 (zygodactyl) may also be reversed. Other possibilities include facultatively reversed digits, and loss of digit 1, or even of digits 1 and 2.
Feathers: Feathers usually (not always) develop in genetically determined patterns along tracts (pterylae). Aves011B&File_type=GIF">BIODIDAC Other areas (apteria) are not feathered. A generalized feather has an unbranched base (calamus) which remains implanted in the feather follicle, (Aves014B&File_type=GIF">BIODIDAC) and an external rachis which supports branches (barbs). The barbs, in turn, may bear barbules which may hook on to the barbules of an adjoining barb. Feathers or portions of feathers with such structures remain compact, flat and sheet-like (pennaceous). Down-like portions without interlocking barbules are referred to as plumaceous. Aves003P&File_type=GIF">BIODIDAC Remiges (see "Wings") and retrices (tail feathers) are almost entirely pennaceous. Body or contour feathers are pennaceous distally. Down is entirely plumaceous with a short rachis. Aves001B&File_type=GIF">BIODIDAC. Numerous specialized variations occur. avian2.html
Ornithology Links: ZOOLOGY 461 ORNITHOLOGY - SPRING SEMESTER 1999; Avian Anatomy ToC (much less cryptic & more useful). ATW020512.
References: Proctor & Lynch (1993)
Checked ATW050717