Evolutionary systematics
Evolutionary Systematics
History
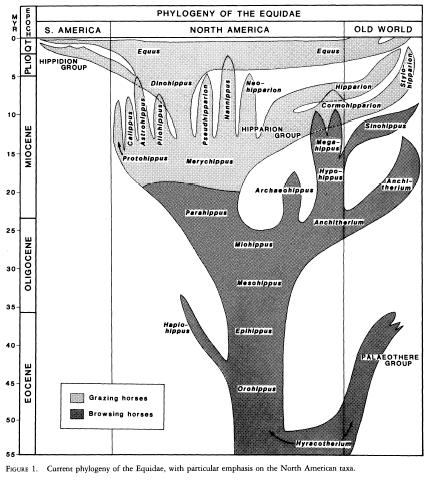
Evolution of the Horse, diagram from Bruce MacFadden, 1985. "Patterns of Phylogeny and Rates of Evolution in Fossil Horses: Hipparions from the Miocene and Pliocene of North America" Paleobiology, Vol. 11, No. 3. (Summer, 1985), pp. 245-257, retrieved from "Laelaps" blog (Brian Switek), The Branching Bush of Horse Evolution. |
Linnaeus, like his 18th century contemporaries, had a static, biblical view of the world. All the species that exist and that he described were the same as those originally created by God, and every species that ever lived was still alive today. This simple worldview was undermined in the late 18th and early 19th century by the discovery of fossil species totally different to anything alive. This led to birth of paleontology, under men like Cuvier and Owen. Cuvier, the father of paleontology, who was the first to name and correctly identify many fossil animals (e.g.: Pterodactylus, Mosasaurus, Didelphys, Palaeotherium) was still a creationist, but explained the existence strange armoured fish, ichthyosaurii, tertiary mammals, mastodons, and the rest in terms of repeated catastrophies, after which God would recreate the world. The biblical flood was considered the most recent of these catastrophes. Owen, who named the order (now superorder) Dinosauria, instead adopted a Goethean concept of evolving archetypes (but not of physical evolution; Owen was strongly opposed to Darwin's theory when it came out). By these sort of mechanisms, Cuvier and Owen could explain the existence of antediluvial (before the flood) monsters. All this changed with Darwin's discovery of the principle of evolution. Darwin, Huxley, and Haeckel established the evolutionary paradigm, and, like Cuvier and Owen, had no problem identifying prehistoric life with Linnaean categories. What evolution did was to make the Linnaean system more dynamic. Thus, Huxley was able to show that Archaeopteryx, the first bird (Class Aves) was also a transitional form between reptiles (Class Reptilia) and modern birds. This synthesis of Darwinian science (specifically the modern evolutionary synthesis) and Linnaean taxonomy was finally fully integrated and systematised in the early 1940s by ornithologist and biological systematist Ernst Mayr, vertebrate paleontologist George Gaylord Simpson, and evolutionary biologist Arthur James Cain. This was the establishment of Systematic Biology, although to distingush it from other schools of biology names like Evolutionary systematics, Evolutionary taxonomy, Evolutionary classification, or Darwinian classification, or Synthetic systematics are used. MAK120229 130331
Methodology
Evolutionary systematics is a way to determine natural relationships of organisms by studying a group in detail and comparing degree of similarity in subspecies, species, and species groups. For example geographical barriers may be sufficient to define subspecies, adaptations to local conditions are features may make a group distinct, and species can be defined as interbreeding local populations. Mayr emphasised geographically determined rings of species, where adjoining races can interbreed, but when the end populations circle back and meet there is geographic isolation. (Cain, 2009 pp.725-6)
As well as considering species, evolutionary systematics also applies to supra-specific taxa, in that groups of species give rise to new groups. Classification reflects both phylogenetic relatedness as well as morphological disparity (overall similarity). The origin of a major new trait or apomorphy (e.g., flowers in angiosperms, endothermy and lactation in mammals) results in the formation of a new "natural group" of the same Linnaean rank as the "natural" group from which it arose (in these examples gymnosperms and reptiles respectively).
In this regard, evolutionary systematics such as palaeontologist Alfred Sherwood Romer, popularised the use bubble or balloon or spindle diagrams that map taxonomic diversity (usually mapped on the horizontal axis) against geological time (mapped vertically, in keeping with the geologists' tendency to equate time with geological strata and hence verticality). The classic example of this, frequently reproduced in old textbooks, is the famous evolution of the horse; see above for a recent version. Thanks to the work of vertebrate paleontologists Romer and Simpson, and later popularisers such as Edwin Colbert, and Romer's student Robert Carroll, evolutionary systematics remained the standard paradigm in paleontology well into the 1990s.
The supremacy of evolutionary systematics in evolutionary theory began to be challenged in the 1960s and 70s by phenetics and especially cladistics, who claimed that it does not have an explicit methodology (much to the surprise of those actually engaged in evolutionary stystematics) or, worse, is "intuitive" (in fact there is no scientific discovery without intuition, as Einstein showed well)
Both Evolutionary systematics and Cladistics use evolution trees, but differ radically in how the tree is drawn. Where each taxon must consist of a single hypothetical ancestor and all its descendants, phylogeny in evolutionary taxonomy allows for groups to be excluded from their parent taxa (e.g. dinosaurs are not considered to include birds, but to have given rise to them). It assumes that ancestor-descendant relationships can be inferred from nodes on phylogenetic trees and considers paraphyletic groups to be natural and discoverable, and at times designated as ancestors (Mayr 1942). Evolutionary systematics also makes possible the organising of organisms into groups (taxa) and hierarchies of such groups (classification systems), in contrast to cladistic, which instead identifies clades and produces cladograms; so both systems can be correct by their own standards. MAK020520 111014 130331
Supraspecific Taxa (Ranks)
It is getting increasingly difficult to find sources which give a balanced comparison of the Linnaean and cladistic methods. Cladistics has simply swept the field. Taxonomy has a good, if somewhat wordy, comparison of the two systems. One of the last, and best, defenses of the Linnaean system -- at least for purposes of nomenclature -- is Benton (2000) which can be accessed here. It would be easy to dismiss these issues as quibbles about nomenclature, but it can make a real difference. The thoughtful student might look briefly at Lane & Benton (2003) . What this paper means, and whether it means anything, depend entirely on on how seriously we take the concept of taxonomic level and exactly how it is defined. Taxonomic level is a concept almost without meaning in a cladistic scheme; while it is critical to the Linnaean view. Lane & Benton (2003) conclude that the shape of the biodiversity curve over time depends on on what taxonomic level is being considered. That issue has important implications in various areas, including public policy. How can we measure diversity without reference to taxonomic level, particularly for systems in which we cannot account for every species? ATW050802.
The decline of evolutionary systematics
With the rise of phenetics and statistical methods, evolutionary systematics was criticised for being based on imprecise, subjective, and complicated sets of rules that only scientists with experience working with their organisms were able to use. It was argued that the resulting phylogenies became impossible to reproduce other than by the specialists themselves, and there was a call for more repeatable and objective methods. Following the short lived career of phenetics, which due to its purely quantifiable approach was unable to distinguish between homology and homoplasy, evolutionary systematics was replaced by cladistics and molecular phylogeny. Because of a misunderstanding between the respective functions and methodologies of evolutionary systematics (concerned with actual phylogenies in deep time) and cladistics (concerned with statistically evaluating different phylogenetic hypotheses) it came to be wrongly believed that they were saying the same thing, that evolutionary systematics is a quantifiable result, and cladograms have to describe the actual evolutionary path of life. Such misinterpretations do disservice to both methodologies, but is surely one of the reasons for the decline and fall of evolutionary systematics. Nevertheless, a number of scientists, such as Tom Cavalier-Smith and the authors of Res Botanica, support evolutionary taxonomy, although their criticism of cladistics only applies to literalist cladistics that mistakes cladograms for actual phylogenies. MAK111014
Why evolutionary systematics still matters
Even today, many diagrams of hominid (= "hominin" evolution are evolutionary systematic and speciation based rather than cladistic and topology-based. This is not because the various homind species can't be represented cladistically - they obviously can - but because it is no longer necessary to do so. As a great many (although cleary not every) species of hominid is now known, there is much less need to posit "hypothetical common ancestors". Instead we are dealing with "actual common ancestors", and with actual phylogeny, a real evolutrionary tree of life, not just phylogenetics and a choice between any number of statistical hypothesis. The same can be said with any well sampled group, such as Carboniferous and Cenozoic foraminifera, Jurassic Ammonite, Neogene bivalves, or Cenozoic mammals. It is not that one is right and the other wrong but that, for example, evolutionary systematics is better with either well known groups and higher linnaean ranks (e.g. ordinal), whereas cladistics does better with poorly known groups sampled on a species (or even individual fossil) level. This is why attempts to claim that only one phylogenetic system is right are by their very nature counterproductive to any attempt to understand the complete (and not just partial) evolutionary history of life on Earth. MAK130331