Bilateria-1
Animals with Bilateral Symmetry
 In general terms, it isn't hard to map out the place of the Bilateria in the scheme of things. The first "animals" were probably uninteresting hollow balls of cells closely related to the living choanoflagellate colonies and -- somewhat more distantly -- to the Fungi. These aggregates were clonal. That is, they all derived from a single parent cell, and might be thought of as a single organism. Since maintaining a hollow ball geometry is trickier than it sounds, the first animals became adept at manipulating cell-cell interactions, which naturally led to the ability to specialize different parts of the "body." After a bit, some deviant globe of cells developed the ability to create new globes and attach them to the original ball. After a few more million years of experimentation, the result was the Porifera, the sponges, which often have no particular shape, but have a respectable degree of specialization within small subunits.
In general terms, it isn't hard to map out the place of the Bilateria in the scheme of things. The first "animals" were probably uninteresting hollow balls of cells closely related to the living choanoflagellate colonies and -- somewhat more distantly -- to the Fungi. These aggregates were clonal. That is, they all derived from a single parent cell, and might be thought of as a single organism. Since maintaining a hollow ball geometry is trickier than it sounds, the first animals became adept at manipulating cell-cell interactions, which naturally led to the ability to specialize different parts of the "body." After a bit, some deviant globe of cells developed the ability to create new globes and attach them to the original ball. After a few more million years of experimentation, the result was the Porifera, the sponges, which often have no particular shape, but have a respectable degree of specialization within small subunits.
The really difficult part came next -- body-level organization. How exactly this was accomplished we won't even guess, at least not right now, since it would be off-topic. The bottom line is that the ball of cells continued to grow, rather than budding new cell aggregates. Since a soft, hollow sphere is not an easy shape to maintain without collapsing, it promptly collapsed. However, it did so in a very interesting and specific way. The maneuver involved creating an inner pocket, as shown in the image. By geometrical necessity, this creates a population of "inside" cells in addition to the original "outside" cells, and a more or less circular boundary region. This is pretty much all we need to create the hydra body plan of a cnidarian a (sea anemone or jellyfish). This is also exactly what happens in the early development of all animal embryos, during the gastrula phase, which is why we have used embryological terminology to label the parts in the diagram from Technau & Scholz 2003).
Ernst Haeckel (1834-1919), whose many accomplishments included looking just exactly like Gandalf the Grey, was the first person to recognize this striking parallel between early animal development and the probable course of early animal evolution. He called his idea the Gastraea Hypothesis, and it is the real source of Haeckel's "Law" that "ontogeny recapitulates phylogeny." This was, and remains, a powerful insight -- true in many cases. It is not a "law" in the same sense as the physical laws of Kepler or Newton. Like many biological "laws," it is, instead, a recurring pattern. It may not apply in any given case, but it happens often enough that it ought to be the default hypothesis until something better comes along. However, in the case of early metazoan evolution, nothing better has come along even now, more than 100 years after Haeckel's time. So, the next time you hear someone belittle Haeckel as it has become fashionable to do), remind them of this remarkable insight. Ask them if they have a better idea.
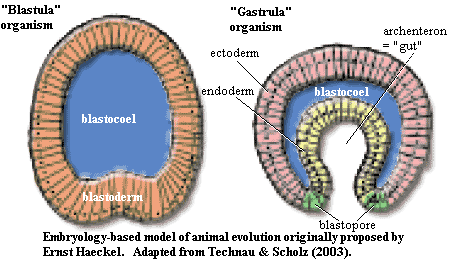 Actually, there are a number of topologically reasonable ways to create an inner compartment, and most can be found within the embryological diversity of the Cnidaria and Ctenophora. All such methods create endoderm and ectoderm, but the gastrula maneuver creates a critical ring-shaped boundary area where ectoderm and endoderm meet. It isn't clear yet why this should have such importance, but it plainly does. In cnidarians, this is the mouth or hypostome. It is the most specialized region of the body, and around it develop the other specialized structures, particularly the tentacles and a large proportion of the sensory cells. This area is clearly a critical link, but extraordinarily hard to understand, as we will discuss later.
Actually, there are a number of topologically reasonable ways to create an inner compartment, and most can be found within the embryological diversity of the Cnidaria and Ctenophora. All such methods create endoderm and ectoderm, but the gastrula maneuver creates a critical ring-shaped boundary area where ectoderm and endoderm meet. It isn't clear yet why this should have such importance, but it plainly does. In cnidarians, this is the mouth or hypostome. It is the most specialized region of the body, and around it develop the other specialized structures, particularly the tentacles and a large proportion of the sensory cells. This area is clearly a critical link, but extraordinarily hard to understand, as we will discuss later.
The hydra body plan, described above, lacks two important elements which are shared by all Bilateria, i.e., Protostomia (bugs, worms, clams, etc.) plus Deuterostomia sea urchins, people, etc.). These were mentioned in the introduction: (1) mesoderm and (2) bilateral symmetry. Mesoderm is a third embryological "germ layer" (fundamental cell type) which develops in the blastocoel after gastrulation. There are several different ways in which mesoderm is produced, which tend to differ between protostomes and deuterostomes. In general, mesoderm formation appears to be based on a key transcription factor coded by the regulatory gene brachyury. We go into this in excessive detail below. The evolution of bilateral symmetry is actually the more difficult development to explain, and we have no easy answers.
The net result of all this reconstruction is a sort of short, worm-like animal with a mouth (but not necessarily an anus), a primitive gut, a specialized area around the mouth with a tendency to develop sensory structures, mesoderm (and, so, probably some internal specializations such as body wall muscles), and bilateral symmetry. That, folks, is Urbilateria.
Urbilateria the ur-bilaterian, the last common ancestor of Attila the Hun and Atta the ant, is a rather critical player in our story because of the way we have defined our clades. As a reminder, the bare bones of our working phylogeny looks like this:
Metazoa (animals) = [toads > toadstools]
├─Porifera (sponges)
└─┬─Cnidaria (anemones, jellyfish, etc.)
└─Bilateria = [men + mollusks] = Urbilateria and all of its descendants
├─Deuterostomia = [men > mollusks]
└─"Middle Earth" (paraphyletic)
└─Protostomia = [bugs + slugs]
├─Ecdysozoa = [bugs > slugs]
└─Lophotrochozoa = [slugs > bugs]
├─Annelida
└─Mollusca
 Bilateria is a crown group. It is defined as the extended family of the last common ancestor of men and mollusks, or Attila and Atta if you prefer. In short, it consists of Urbilateria and all of its descendants. Since it is a crown group, it has two branches, one leading to men and the other leading to mollusks. Deuterostomia is a stem group, defined to take in everything more closely related to men than mollusks. That is, it takes up all of the branch of Bilateria leading toward humans. However Protostomia is not a stem group. It is another crown group, defined by the intersection of bugs (Ecdysozoa) and slugs (Lophotrochozoa). Therefore, there is a gap of unknown size and composition between Urbilateria and Protostomia containing animals which are bilaterians, but neither deuterostome nor protostome. We have labeled this part of phylospace "Middle Earth," since it may be completely mythical, or quite real and filled with all manner of unfamiliar creatures. In order to get a handle on Bilateria, the challenge is to find out if Middle Earth exists and, if so, who lives there. That, in turn, may depend on what characteristics Urbilateria might have had.
Bilateria is a crown group. It is defined as the extended family of the last common ancestor of men and mollusks, or Attila and Atta if you prefer. In short, it consists of Urbilateria and all of its descendants. Since it is a crown group, it has two branches, one leading to men and the other leading to mollusks. Deuterostomia is a stem group, defined to take in everything more closely related to men than mollusks. That is, it takes up all of the branch of Bilateria leading toward humans. However Protostomia is not a stem group. It is another crown group, defined by the intersection of bugs (Ecdysozoa) and slugs (Lophotrochozoa). Therefore, there is a gap of unknown size and composition between Urbilateria and Protostomia containing animals which are bilaterians, but neither deuterostome nor protostome. We have labeled this part of phylospace "Middle Earth," since it may be completely mythical, or quite real and filled with all manner of unfamiliar creatures. In order to get a handle on Bilateria, the challenge is to find out if Middle Earth exists and, if so, who lives there. That, in turn, may depend on what characteristics Urbilateria might have had.
We will use two distinct approaches to investigate the population of Middle Earth: phylogeny and embryology. The phylogenetic approach involves the usual business of comparing warring cladograms and methodologies. This approach focuses on the point where bugs and slugs diverge. Anything bilaterian, but basal to that point, is a hobbit. The embryological approach, by contrast, focuses on Urbilateria. It tends to be difficult, theoretical and often quite speculative. However, as we will see, there are excellent reasons for resorting to this sort of thing, as a check on the usual phylogenetic methods.
Trees in the Entwood
 We would love to say that no one could possibly review all of the recent literature on bilaterian phylogeny. Unfortunately several inconsiderate show-offs evidently had nothing better to do, and have published just such reviews, too recently to ignore. Zrzavý 2001); Halanych (2004), Philippe et al. (2005). Confronted with the undeniable evidence of all this industry, we can claim only that no one as shiftless as we are could possibly review all of the recent literature on bilaterian phylogeny. Nonetheless, we have dabbled in the literature in our usual cursory manner -- at least to the extent of casually skimming the said reviews of Zrzavý, Halanych, and Philippe et al.
We would love to say that no one could possibly review all of the recent literature on bilaterian phylogeny. Unfortunately several inconsiderate show-offs evidently had nothing better to do, and have published just such reviews, too recently to ignore. Zrzavý 2001); Halanych (2004), Philippe et al. (2005). Confronted with the undeniable evidence of all this industry, we can claim only that no one as shiftless as we are could possibly review all of the recent literature on bilaterian phylogeny. Nonetheless, we have dabbled in the literature in our usual cursory manner -- at least to the extent of casually skimming the said reviews of Zrzavý, Halanych, and Philippe et al.
Fortunately, a few examples will suffice to make the relevant point. What we will find is that, while we can obtain some hints, the trees of Middle Earth often turn out to be Ents. That is, they are not fixed in place, but shift their limbs in unexpected ways, with a tendency to ensnare unwary travelers with their long branches. In short, these trees are unstable and exhibit long branch attraction.
Consider Steinauer et al. 2005). One of the problems with investigating Middle Earth is that its inhabitants are poorly known. Most candidate species are rare and/or obscure parasites. The Acanthocephala are a case in point. The acanthocephalan Leptorhynchoides is a parasite of vertebrate carnivores which looks a little like a nemertine worm with training wheels. Virtually the only persons with an interest in Leptorhynchoides are a few, notoriously single-minded, public health types whose interest begins and ends with finding new methods for making the Acanthocephela yet rarer and/or more obscure.
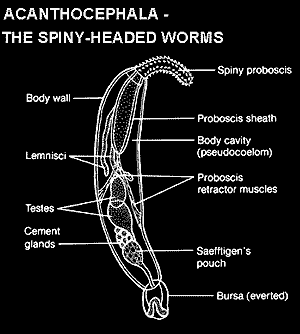 To the extent that anyone has cared at all, the understanding has been that acanthocephalans are some kind of evil rotifer cousin (Herlyn et al., 2003) or just some kind of evil rotifer (Zrzavý, 2001). Undeterred by considerable morphological evidence, Steinauer et al. sequenced the entire mitochondrial genome of Leptorhynchoides. They found many useful bits of information which, as in most mtDNA studies, they proceeded to ignore completely (see methodological critique at Insectivora). These workers then applied the usual sequence-based techniques to construct a phylogenetic tree. Perversely, they failed to include any sequences from rotifers, gnathostomulids, chaetognaths, or, in fact, anything that might actually be comparable to Acanthocephala. They found that (a) nothing lives in Middle Earth; (b) acanthocephalans are close to Platyhelminthes; (c) both are mollusks; and (d) that long branch attraction might have played a part in the results.
To the extent that anyone has cared at all, the understanding has been that acanthocephalans are some kind of evil rotifer cousin (Herlyn et al., 2003) or just some kind of evil rotifer (Zrzavý, 2001). Undeterred by considerable morphological evidence, Steinauer et al. sequenced the entire mitochondrial genome of Leptorhynchoides. They found many useful bits of information which, as in most mtDNA studies, they proceeded to ignore completely (see methodological critique at Insectivora). These workers then applied the usual sequence-based techniques to construct a phylogenetic tree. Perversely, they failed to include any sequences from rotifers, gnathostomulids, chaetognaths, or, in fact, anything that might actually be comparable to Acanthocephala. They found that (a) nothing lives in Middle Earth; (b) acanthocephalans are close to Platyhelminthes; (c) both are mollusks; and (d) that long branch attraction might have played a part in the results.
We can certainly endorse the last conclusion. If a group of DNA sequences are similar, one can sometimes create a valid phylogenetic tree by comparing the changes at homologous sites. Over time, however, the same site will have changed multiple times. The nucleotide found at the site becomes randomized and carries no valid phylogenetic signal. See discussion at Insectivora. The relevant time frame varies with the site, the species, and by blind luck. Consequently, when one is comparing sequences which diverged long ago and are quite different ("long branches"), the phylogenetic signal is swamped by random noise and by other, non-phylogenetic, effects, giving rise to all kinds of spurious results. This is called "long branch attraction." Unfortunately, long branch attraction actually becomes worse as more data are added to the matrix. Philippe et al. (2005). This effect may be analogous to a subtle problem related to missing data in morphological work. See discussion at Pythonomorpha. However, contrary to a wrong, but oft-repeated, scientific fable, morphological parsimony methods are generally much less subject to this source of error. Siddall & Whiting (1998).
For reasons we really ought to discuss one day, small, parasitic organisms tend to mutate a lot faster than others. Most of the possible inhabitants of Middle Earth are just such organisms and are known to mutate faster than a comic book superhero. Lavrov & Lang (2005) (nematode mtDNA "statistically indistinguishable from randomly shuffled genomes with the same gene content") Thus these organisms quickly reach branch lengths at which any phylogenetic signal is overwhelmed by noise.
 Treehugging
Treehugging
In the last few years, a number of groups have attempted various strategies to confine these phylogenetic Ents. While they have had somewhat more success than Sauruman the White, the results have been unsatisfactory.
Lavrov & Lang (2005) used the promising technique of comparing mitochondrial gene order (not sequence); but they were forced to eliminate taxa with unacceptably high rates of branch growth. They obtained believable results, but resolution was poor and all of the likely inhabitants of Middle Earth had to be pruned to make their approach work. Much the same might be said of Philippe et al. (2005), who use 18S rRNA sequences. Both recognize the long-branch problem, attempt to deal with it, and recover reasonable trees -- but with very little phylogenetic resolution and no information on the phyla of interest to us.
A more productive approach may be that of Ruiz-Trillo et al. (2002), who used one of the myosin genes to map bilaterian phylogeny. Myosin is one of the critical contractile proteins of all metazoan cells -- and almost all eukaryotic cells. The reasoning, which seems to have been sound, was that myosin is under more or less uniform structural constraints and ought to evolve both slowly and regularly. The authors conclude that acoel worms and nemertodermatids (collectively, the Acoelomorpha) are basal to the rest of the former Platyhelminthes which are protostomes. The only difficulty is that the Acoelomorpha also end up basal to the crown group Bilateria! As it turns out, this is entirely possible. See discussion of Cook et al. (2004), toward the end of all this noise. Nevertheless, Ruiz-Trillo et al. (2002) requires no panic-driven redefinition of the Bilateria. We certainly thought about it, but soon stopped screaming (when we ran out of breath). Restored to our customary state of cheerful pomposity, we set about the serious business of making excuses. Fortunately, the branch order of a number of the major clades seems to be slippery in this study. Accordingly, we can't take it as solid authority that Acoelomorpha has fallen right out of Bilateria.
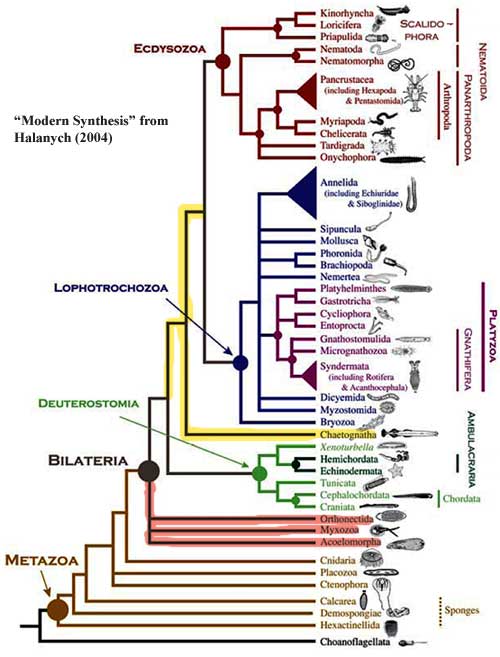 Halanych 2004) has attempted to summarize those results in the figure reproduced here. However, it is unclear what definition he is applying to Bilateria. In our scheme, there are only three kinds of Bilateria: deuterostomes, protostomes and hobbits (stem protostomes -- the yellow lines in the figure). Since Halanych supplies no phylogenetic definition, we were forced against our will, and with much gnashing of teeth, rending or garments, etc., to consult the primary sources for the three groups marked with red lines.
Halanych 2004) has attempted to summarize those results in the figure reproduced here. However, it is unclear what definition he is applying to Bilateria. In our scheme, there are only three kinds of Bilateria: deuterostomes, protostomes and hobbits (stem protostomes -- the yellow lines in the figure). Since Halanych supplies no phylogenetic definition, we were forced against our will, and with much gnashing of teeth, rending or garments, etc., to consult the primary sources for the three groups marked with red lines.
As it turns out, the Orthonectida are almost certainly metazoans, particularly under our stem group definition; but it's anyone's guess whether they, or any other members of the mysterious Mesozoa, are also bilaterians. Hanelt et al. (1996). At any rate neither Hanelt's study nor any later work exclude the Mesozoa from Middle Earth. Zrzavý 2001).
The Myxozoa may well reside in Middle Earth. Monteiro et al. (2002). Zrzavý 2001) asserts that myxozoans are more primitive, but his review was written before Monteiro's report that the morphologically bilaterian Buddenbrockia is a myxozoan.
The Acoelomorpha are consist of two different phylum-level groups, the Nemertodermatida and the Acoela. Both of these groups were parts of the classical Phylum Platyhelminthes. However, molecular studies consistently separate the acoelomorphs, and the separation is plausible on morphological grounds. We will return to these candidate hobbits later. For the moment, it is enough to note that both are likely bilaterians, but either or both may be protostomes.
Finally, Halanych places the Chaetognatha right in the middle of Middle Earth. This is recent work from Halanych's own lab, and it has not yet attracted much support from elsewhere. Chaetognaths ("arrow worms") were considered to be deuterostomes until quite recently, when they were relocated to the protostomes. Possibly the thought of moving this lot yet again has inspired some resistance.
"It is a lovely language, but it takes a very long time to say anything in it."
Phylogenetic methods may ultimately be able to sort out the long branch problem by using carefully chosen, highly conserved sequences. But phylogenetic methods, particularly molecular methods, can also suffer from an opposite problem, i.e. not just long branches, but short stems. A branching pattern can best be resolved if the radiation occurs in a relatively slow and orderly way, so that each critical divergence is clearly separate from the previous one. However, Middle Earth was being populated right in the midst of the Cambrian Explosion -- arguably the most rapid series of evolutionary changes which the Animal world has ever experienced.
The Molecule Mob have always argued that the Cambrian Explosion did not exist, and that the actual divergences occurred deep in the Neoproterozoic. However, if that were the case, as some of the Moleculons now admit, these tree-building methods ought to work much better than they in fact do. Rokas et al. (2003).
Indeed, Rokas et al. (2005) have now turned the argument on its head. They make a fairly convincing case that the failure of sequence-based methods to reconstruct the tree at this point in time tends to prove the existence of a Cambrian Explosion. They point out that, if the Explosion did occur, and if it lasted significantly less than 50 My (which is a reasonable bet on morphological grounds), even the complete sequence of the all living lineages from that event may be mathematically insufficient to reconstruct the branching pattern. In other words, it may not even theoretically possible to construct the correct tree using these methods. The fact that we cannot seem to get consistent results using these methods strongly suggests that the metazoan phyla in fact diverged within a short space of time.
Note, however, that Levinton et al. (2004), using the same theoretical framework, claim that that they have recovered the correct phylogeny using sequence methods, so that the Cambrian Explosion cannot have existed. However, the phylogeny they rely on (Wray et al., 1996) actually draws a tree of only 6 metataxa, using individual taxa and methods which are not fully described. We view this dispute with placid ambivalence, suspecting that the truth lies between the positions garrisoned by these contending forces.
page uploaded MAK020407, last modified ATW050724
checked ATW050724
this material may be freely used as long as attribution is given