Bilateria-2
"Where many paths and errands meet"
 Given the very serious problems with the usual phylogenetic methods, we are forced back onto embryology. Unfortunately, although Haeckel's dictum holds remarkably well in getting us from choanoflagellates to Cnidaria, it makes no robust predictions about Urbilateria or Middle Earth. Nevertheless, by triangulating through the known developmental scripts of existing organisms, it has been possible to say a few useful things about Urbilateria and the inhabitants of Middle Earth. In a few cases, we can look to classical developmental biology, as Haeckel did. However, in most cases, we must depend on the homology of transcription factors, the proteins that control developmental programs by promoting or suppressing the expression of particular genes.
Given the very serious problems with the usual phylogenetic methods, we are forced back onto embryology. Unfortunately, although Haeckel's dictum holds remarkably well in getting us from choanoflagellates to Cnidaria, it makes no robust predictions about Urbilateria or Middle Earth. Nevertheless, by triangulating through the known developmental scripts of existing organisms, it has been possible to say a few useful things about Urbilateria and the inhabitants of Middle Earth. In a few cases, we can look to classical developmental biology, as Haeckel did. However, in most cases, we must depend on the homology of transcription factors, the proteins that control developmental programs by promoting or suppressing the expression of particular genes.
Reading the Runes
Runic scripts are writing schemes which tend to fall in between pictograms and context-dependent constructs like alphabets. Erwin & Davidson (2002) remind us that transcription factors are somewhat the same way. They argue that just because (1) many protostomes and deuterostomes have eyes and (2) the pax-6 transcription factor is involved in the morphogenesis of eyes in both cases does not mean that the eyes of protostomes and deuterostomes are homologous, or even that Urbilateria had eyes. The lowest common denominator is pax-6 regulation of photosensitive pigments. These pigments may be found in eyes in many organisms, but they may also have been spread throughout the ectoderm of Urbilateria, for all we know, as they are in some echinoderms. On this basis, Erwin & Davidson conclude that Urbilateria was probably a very simple organism, possibly similar to the Ediacaran Kimberella (or the extant, and even simpler, Xenoturbella).
It's a good point. The meaning of these runes depends on context and cannot be equated one-to-one with some particular morphological expression. But they aren't a completely arbitrary alphabet, either; and, besides, we know something about their context. Like most questions of homology, it is not entirely a yes-or-no affair. The eyes of protostomes and deuterostomes are homologous -- to the extent that they both use the pax-6 regulatory system in a way which seems to have been unchanged since their last common ancestor (by definition, Urbilateria). But, whether we can make stronger claims to homology depends on what other evidence we have. Consider a few examples.
 The Underworld of Middle Earth
The Underworld of Middle Earth
Perhaps the most fundamental issue relates to the formation of mesoderm. The single most basic difference between bilaterians and all other animals is the presence of this third germ layer. If mesoderm is not homologous in all bilaterians, then we may as well pack it in and quit blathering about Bilateria, because Bilateria won't be a monophyletic group.
Unfortunately, we aren't going to be able to weasel out of this discussion so easily. Technau & Scholz 2003) have recently reviewed this area. Here, the case for homology is strong, and we may be reasonably certain that all hobbits have mesoderm. To put the matter briefly, mesoderm formation is run by some combination of the following four genes (or gene families): twist, snail, brachyury, and the GATA family. Despite the names, this is probably not a crime syndicate. The GATA family, for example, is supposedly named for part of the sequence to which the GATA gene products bind. On the other hand, the GATA family are elusive, hard to pin down, and appear to operate by "facilitating" the business of higher level inducers, "enforcing" their directives at the level of individual genes. So, it's safer to be ... respectful.
The GATAs are unique to Bilateria and may be a synapomorphy of the clade. The other factors play rather interesting roles in the Cnidaria. Twist is responsible for muscle, or, rather it "induces" endodermal cells to form muscle. The endoderm might make muscle anyway, if asked politely, but twist is a very convincing inducer. It hangs around just in case, to keep everyone ... respectful. Snail is also involved in muscle formation, but its role lies in making the right contacts -- regulating cell-to-cell adhesion or mobility, making the right arrangements. Brachyury just sits on the lip of blastopore. It doesn't seem to do much of anything, except to offer little suggestions to cells migrating into the archenteron. Mostly those cells are, as we said, respectful. But, if not, it turns out that brachyury happens to be very close friends with twist and snail.
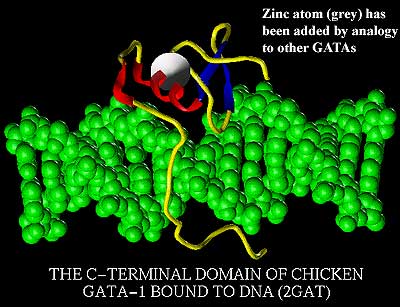 Muscle is one of the basic mesodermal tissues in bilaterians, and it is easy to see how this very efficient arrangement could lead to the formation of an entirely new germ layer. Rieger & Ladurner (2003). All that is needed is to recruit more lieutenants to take on more specialized jobs, which is presumably where the GATA Family came in. While this is an elegant explanation of mesoderm evolution, it seems that not everyone agrees. See, e.g., Lartillot et al. (2002), reporting that Patella orthologs of twist and snail are not involved in mesoderm specification. Perhaps so, but we are unconvinced. Mind you, this has absolutely nothing to do with the alleged little gift we allegedly received from the alleged GATA brothers. They don't even exist. It's all malicious gossip. Honest! We just believe in being ... respectful.
Muscle is one of the basic mesodermal tissues in bilaterians, and it is easy to see how this very efficient arrangement could lead to the formation of an entirely new germ layer. Rieger & Ladurner (2003). All that is needed is to recruit more lieutenants to take on more specialized jobs, which is presumably where the GATA Family came in. While this is an elegant explanation of mesoderm evolution, it seems that not everyone agrees. See, e.g., Lartillot et al. (2002), reporting that Patella orthologs of twist and snail are not involved in mesoderm specification. Perhaps so, but we are unconvinced. Mind you, this has absolutely nothing to do with the alleged little gift we allegedly received from the alleged GATA brothers. They don't even exist. It's all malicious gossip. Honest! We just believe in being ... respectful.
Rings of Power
Another point which now seems secure is that Urbilateria was not segmented. Seaver (2003). Segmentation is best understood in arthropods and, more generally, in the Ecdysozoa. Almost all ecdysozoans are segmented but, tellingly, their most basal living group, the nematodes, are not segmented. In insects, segmentation involves two types of transcription factors: polarity genes particularly engrailed) and pair-rule genes. While polarity genes are quite commonly used in the same way in other ecdysozoans, the pair-rule genes are not. They appear to play a secondary role. Seaver (2003).
Most lophotrochozoans are unsegmented. Among the segmented annelids, the segmentation pattern is established through stereotyped spiral divisions in early embryogenesis, which is unlike the insect model. The initiating embryological tissue varies a good bit within the Annelida, and even between adult and larval segments. Like vertebrates, but unlike insects, segmentation proceeds in an anterior to posterior sequence; and; unlike both, the number of segments is not even approximately fixed, and 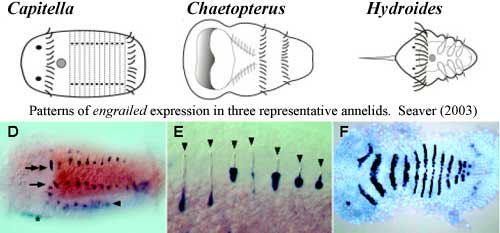 segments continue to develop after embryogenesis is complete. Engrailed is involved in segmentation, but it seems to be a secondary messenger. The pattern of engrailed expression differs markedly even among the Polychaeta, and normal development is possible after ablation of engrailed-expressing cells. The heavy lifting is done, instead, by a completely different set of genes, the "gap" genes, which have no role in insect segmentation. Seaver (2003)
segments continue to develop after embryogenesis is complete. Engrailed is involved in segmentation, but it seems to be a secondary messenger. The pattern of engrailed expression differs markedly even among the Polychaeta, and normal development is possible after ablation of engrailed-expressing cells. The heavy lifting is done, instead, by a completely different set of genes, the "gap" genes, which have no role in insect segmentation. Seaver (2003)
There is no evidence that any of the basal deuterostomes were segmented. Segmentation was re-invented among early chordates. Unlike any protostome, chordates use a rather delicate timing mechanism based on oscillating expression of the notch gene to produce segments during elongation of the dorsal midline. This creates mesodermal aggregates, somites, which actually create the segmental structures. Pair rule, polarity and gap genes are present, but seem to have little or no role in segmentation. Accordingly, we can be fairly confident that Urbilateria was not segmented and that any hobbits, orcs, elves, etc. are unlikely to exhibit segmentation. Seaver (2003).
The Eyes of Sauron
As discussed, Erwin & Davidson (2002) use pax-6 and eye development as an example of the crawling evils of naive reliance on crude transcription factor homologies. Actually, a more current version of this story, as reviewed by Arendt & Wittbrodt (2001), is a rather more urbane and polished tale. Arendt and co-workers have further refined the work since that time. Fortunately, it is not necessary for us to summarize this work, since Pharyngula has conveniently predigested the data at Rhabdomeric and ciliary eyes. A lot more than pax6 backs up the claim that eyes have extensive homologies across Bilateria.
 For those too indifferent even to read Pharyngula's abbreviated account, here's the TV Guide version:
For those too indifferent even to read Pharyngula's abbreviated account, here's the TV Guide version:
Light comedy. Two photosensitive young systems, the ciliary and rhabdomeric, meet in the Proterozoic where they both work as photoperiod detectors. In the deuterostomes, they move in together, but the ciliary system starts seeing others, and the rhabdomeric system is relegated to a supporting role and menial household work. In the protostomes, they break up, and the rhabdomeric system goes on to have a glamorous career in pictures while the ciliary cells have a dead-end job in the brain. Cast of billions. 600 My, mostly B&W or limited Color, PG-13 (adult and/or larval situations, some nudibranchs).
"Hobbits ... are inclined to be fat in the stomach"
This section could well be called Haeckel's revenge. A number of workers have recently obtained spectacular results with old-fashioned descriptive embryology. In part, the quality of these papers is due to the availability of sophisticated molecular probes for looking at gene expression. However, some of the improved quality simply results from doing better embryology. The tendency in the past has been to compare embryos of vertebrates and Drosophila. But both have very specialized developmental patterns and are unlikely to tell us much about Urbilateria.
The tale from the vertebrate-fruit fly comparison is well known. In protostomes, the mouth develops "first." More accurately, the blastopore elongates along the future ventral midline and the mouth and anus develop at opposite ends. In deuterostomes, the gut elongates. The blastopore forms only the anus, and the mouth is formed by punching a new hole in the ectoderm along the future ventral midline. This seems very different only if one bases the comparison on vertebrates, in which mouth formation is delayed and involves some very peculiar behavior by the surrounding ectoderm. See, e.g., The Basisphenoid and Teeth: Overview.
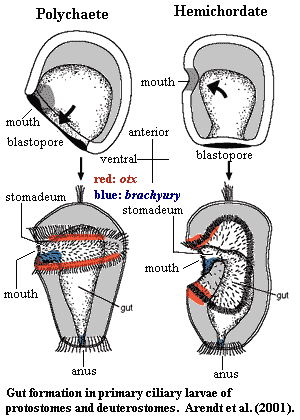
Arendt et al. (2001) do the sensible thing and compare the surprisingly similar primary ciliated larvae of polychaete annelids and hemichordates. Despite the fact that these are protostome and deuterostome, respectively, their primary larvae are strikingly similar in morphology and, as it turns out, in the way they express the key transcription factors related to gut formation: otx, brachyury and goosecoid. At this point, we are moved to remind the enthusiastic reader of the teachings of Erwin & Davidson (2002). These factors may not have the same degree of priority in the developmental scheme. They may not be doing the same thing -- but it's a very pretty picture all the same.
We lack the space for a full description of what Arendt & Co. did, but the bottom line is a strong case for homology of a three- (actually four-) part gut with a mouth, stomadeum (pharyngeal region in vertebrates), an undifferentiated middle section, and an anus.
The Wisdom of the Elves
 Last, but far from least, we consider the information on the nervous system; and, indeed, the subject makes us nervous. We have readily confessed that the scope of this essay has forced us to rely on secondary sources. Here, we have been schooled by Alain Ghysen 2003) who produces utterly intimidating work at the intersection of molecular, developmental, evolutionary and neurobiology at the Université Montpellier II, when he is not raising orchids or studying poetry in an interesting array of archaic languages. Whether Prof. Ghysen is actually an elf, we cannot say. Before rejecting this hypothesis, consider also that he is a dedicated archer. We can say that he, like an elf, is sometimes opinionated and, when asked to give his opinion, he is rarely shy about expressing his views. In Ghysen (2003), he gives fair warning that he will "venture into somewhat speculative grounds."
Last, but far from least, we consider the information on the nervous system; and, indeed, the subject makes us nervous. We have readily confessed that the scope of this essay has forced us to rely on secondary sources. Here, we have been schooled by Alain Ghysen 2003) who produces utterly intimidating work at the intersection of molecular, developmental, evolutionary and neurobiology at the Université Montpellier II, when he is not raising orchids or studying poetry in an interesting array of archaic languages. Whether Prof. Ghysen is actually an elf, we cannot say. Before rejecting this hypothesis, consider also that he is a dedicated archer. We can say that he, like an elf, is sometimes opinionated and, when asked to give his opinion, he is rarely shy about expressing his views. In Ghysen (2003), he gives fair warning that he will "venture into somewhat speculative grounds."
He then proceeds to erect a strongly fortified position around the proposition that Bilateria not only had a reasonably well-organized nervous system, but also the framework of a multi-part brain, brainstem, longitudinal nerve cord, and several types of specialized sensory cells, particularly mechanosesnsory cells. The bit about the proto-brain sounds like a stretch, but others, working from wholly different premises, have come to the same conclusion. Meinhardt 2002) (mainly body patterning in early development). Ghysen, as one might expect, relies on a battery of molecular, developmental, evolutionary, etc. evidence. We won't attempt to summarize it here, since we have gone on too long as it is. At least that's our story, and we're sticking to it. In fact, it is already time to move on to the next section:
page uploaded MAK020407, last modified ATW050724
checked ATW050724
this material may be freely used as long as attribution is given