| Palaeos: |
|
Rhizaria |
| EUKARYA | Rhizaria |
| Page Back | Unit Back | Unit Home | References | Glossary | Pieces |
| Page Next | Unit Next | Life | Dendrogram | Taxon Index | Time |
Eukarya |--Metamonada `--+--Discicristata `--+--RHIZARIA | |--Radiolaria | | |--Acantharia | | `--Polycystinea | `--+--+--Acetosporea | | `--+--Xenophyophorea | | `--Foraminifera | `--+--Phytomyxea | `--Cercozoa `--+--+--Chromalveolata | | |--Alveolata | | `--Chromista | `--Plantae `--Stem Metazoa |--Fungi `--Metazoa |
Introduction Phylogeny Diversity Radiolaria Acantharia Polycystinea Acetosporea Foraminifera Phytomyxea Cercozoa References
|
 The clade Rhizaria of unicellular eukaryotes was named very recently (Cavalier-Smith,
2002), but has rapidly ingratiated itself as an industry
standard. It contains a large number of mostly amoeboid organisms,
including such significant groups as the radiolarians and
foraminiferans.
The clade Rhizaria of unicellular eukaryotes was named very recently (Cavalier-Smith,
2002), but has rapidly ingratiated itself as an industry
standard. It contains a large number of mostly amoeboid organisms,
including such significant groups as the radiolarians and
foraminiferans.
So far, Rhizaria seems to be supported solely by molecular data – there are no morphological characters unique to the clade. Most are biciliate amoeboflagellates, at least at some point in the life cycle – though many have dispensed with flagella altogether. Pseudopodia are root-like reticulopodia, filopodia and/or axopodia – not broad lobopodia as in Amoeba. All of these features can, however, be found in members of other clades. Nevertheless, the Rhizaria are supported by both rRNA and actin trees (Cavalier-Smith & Chao, 2003; Nikolaev et al. 2004), and are probably here to stay.
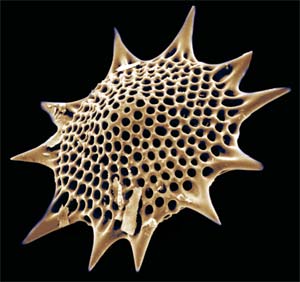
Image: unidentified radiolarian from the Hamilton College Electron Microscopy site.
© Christopher Taylor 2004. CT041217
Rhizaria
|-—Radiolaria
| |--Polycystinea
| `--+--Sticholonche
| `--Acantharea
`--+--+--Gromiidae
| `--+--Ascetosporea
| `--Foraminifera
`--+--Phytomyxea
`--+--Desmothoracida
`--‘core Cercozoa’ (including Phaeodarea)
|
SSU rRNA and actin trees both give a similar picture of rhizarian phylogeny – the phylogeny below is derived from Nikolaev et al. (2004) and Polet et al. (2004). The positions of Sticholonche and Ascetosporea, however, should be treated with some suspicion.
Cavalier-Smith (2002; Cavalier-Smith & Chao, 2003) previously suggested a monophyletic clade, Retaria, formed by Radiolaria and Foraminifera, characterised by reticulose pseudopods in both groups, and supported weakly by molecular phylogenies. More recent analyses fail to support this grouping, and reticulopodia probably evolved independently in the two groups – an adaptation to large size in both? Instead, Foraminifera group with Gromia, a marine amoeboid with smooth filopodia that produces an organic test like many basal Foraminifera (Longet et al. 2003).
© Christopher Taylor 2004. CT041217
The Rhizaria can be thought of as being composed of Radiolaria, Foraminifera, and Cercozoa. That's fortunate, since we are going to treat Rhizaria in that very manner.
Radiolaria
|
||
| Radiolarians |
Polycystinea
|
|
Acantharea
|
||
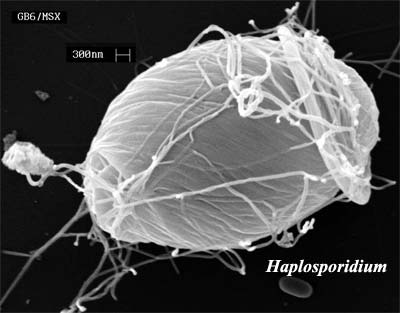
The foraminiferan clades are highly contentious, which is too bad. The evolution and diversity of these sturdy, testate Eukarya form an important part of Mesozoic history and stratigraphy. A better understanding of their ?Cambrian beginnings might give us a bater handle on their later development. We treat only one group of stem Foraminifera here, the Acetosporea.
Acetosporea
|
|
Foraminifera
|
Finally, the cercozoan group:
Phytomyxea
|
|
Cercozoa
|
| Page Back | Page Top | Unit Home | Page Next |