| Palaeos: |
|
Plantae |
| EUKARYA | Plantae |
| Page Back | Unit Back | Unit Home | References | Glossary | Pieces |
| Page Next | Unit Next | Life | Dendrogram | Taxon Index | Time |
Eukarya |--Metamonada `--+--Discicristata `--+--Rhizaria `--+--+--+--Alveolata | | `--Chromista | `--PLANTAE | |--Glaucophyta | `--+--Rhodophyta | `--Chlorobionta `--Stem Metazoa |--Fungi `--Metazoa |
Introduction Plant Phylogeny Plant Diversity Glaucophyta Rhodophyta Chlorobionta References
|
 This
section grows out some comments on the main plant section, now called Chlorobionta
(land plants and green algae) from Chris Taylor, one of our regular
contributors. He objected that the "plants" go deeper than that,
to include the Glaucophyta and red algae (Rhodophyta). As usual, he was
correct. Fortunately, having used the term Chlorobionta for land
plants and green algae, we had accidentally freed up the vaguer
"Plantae" to use for a more inclusive group.
This
section grows out some comments on the main plant section, now called Chlorobionta
(land plants and green algae) from Chris Taylor, one of our regular
contributors. He objected that the "plants" go deeper than that,
to include the Glaucophyta and red algae (Rhodophyta). As usual, he was
correct. Fortunately, having used the term Chlorobionta for land
plants and green algae, we had accidentally freed up the vaguer
"Plantae" to use for a more inclusive group.
Its hard to give Plantae a reasonable phylogenetic definition. There are three, and possibly four, living groups which diverge from the base of the Plantae: the red algae, the glaucophytes, and the green plants (plus green algae). Just possibly, the Cyanidiales, usually placed at the base of the red algae, represent a fourth basal branch. But see, e.g., Ciniglia et al. (2004). All of these groups share the incorporation of a cyanobacterium as an organelle, i.e., a chloroplast. There is cautious general agreement that this critical bit of indigestion probably happened only once in this group, so that all of the Plantae descend from a single common ancestor. What we don't know is the nature of the beast before it started cultivating house plants.
We also are very uncertain what the sister group of the Plantae might be. Our working hypothesis is that it is the Alveolata + Chromista group. However, many would disagree, and it is not a strongly-supported hypothesis. One significant cause of difficulty is the bizarre origin of the chloroplasts of Chromista. It appears that, just as some ancestral plant first acquired a chloroplast by failing to digest a cyanobacterium, the ancestral chromist acquired a chloroplast by failing to digest a plant. The resulting hybrid organism, with its potential for three-way lateral gene transfer, seems to have been designed by some malicious deity with the specific intent to make accurate phylogenetic analysis impossible.
So, who are we to quarrel with divine providence? Bowing to the ineffable, we will, for the moment at least, leave Plantae as "things with primary chloroplasts." This kind of apomorphy-based definition almost always leads to grief in the long run, but we have no good alternatives.
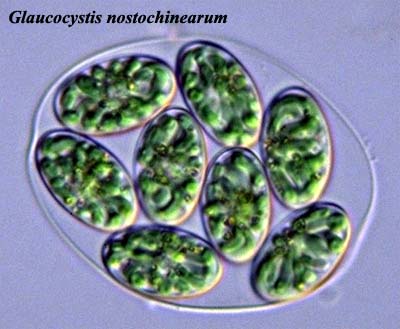
Image: Glaucocystis from the Protist Information Server.
ATW050128. Text public domain. No rights reserved.
 The basal phylogeny of plants is simplicity itself, largely because very
little survives near the base of the Plantae. Undoubtedly, a good many
interesting plant types evolved in the 1-3 Gy since the first plant acquired its
chloroplast. However the fossil record is essentially non-existent for all
but a very few types.
What we have today are the Glaucophyta, Rhodophyta, and Chlorobionta. Of
these three, the glaucophytes are plainly the most unspecialized. As the
image of Glaucocystis shows, this is a rather plain vanilla organism,
largely a sack full of chloroplasts. The chloroplasts themselves are also
primitive. They retain a "cell wall" of peptidoglycan
which ought to dispel any doubts about their bacterial origin. Further,
the thylakoid
membranes of the chloroplasts are not stacked, suggesting a
significantly different, and perhaps more primitive, evolutionary path.
The basal phylogeny of plants is simplicity itself, largely because very
little survives near the base of the Plantae. Undoubtedly, a good many
interesting plant types evolved in the 1-3 Gy since the first plant acquired its
chloroplast. However the fossil record is essentially non-existent for all
but a very few types.
What we have today are the Glaucophyta, Rhodophyta, and Chlorobionta. Of
these three, the glaucophytes are plainly the most unspecialized. As the
image of Glaucocystis shows, this is a rather plain vanilla organism,
largely a sack full of chloroplasts. The chloroplasts themselves are also
primitive. They retain a "cell wall" of peptidoglycan
which ought to dispel any doubts about their bacterial origin. Further,
the thylakoid
membranes of the chloroplasts are not stacked, suggesting a
significantly different, and perhaps more primitive, evolutionary path.
Likely prasinophyte (basal chlorobiont) remains are known from close to 1500 Mya. Javaux et al. (2004). If these have been correctly identified, the division between red and green algae must also be extremely ancient. However, the two algal groups have enough in common to be virtually certain that their divergence post-dates the origin of the Glaucophyta.
Image: Microcladia from the UCLA OceanGLOBE site.
ATW050131. Text public domain. No rights reserved.
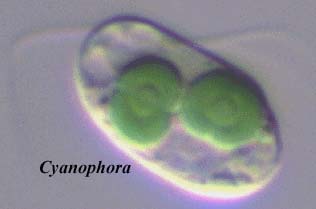 This group is also known as the Glaucocystophyta, for the reason that the
type genus is Glaucocystis -- to which we respond, "so
what?" The type species of the Rhodophyta is probably not Rhodis,
or algal taxonomy is in very deep trouble. Rhodis is the
rhododendron, and not known to be planktonic. Accordingly, we will retain the traditional name for this little
taxon of three genera: Glaucocystis, Cyanophora, and Gloeochaete. Other
genera have been included, at times, based largely on the presence of cyanelles
(primitive chloroplasts). However these appear, on closer inspection, to be
unrelated. In particular, Paulinella is not a glaucophyte, but an
amoeba which apparently acquired its cyanelle by independent primary
endosymbiosis. McFadden
(2001).
This group is also known as the Glaucocystophyta, for the reason that the
type genus is Glaucocystis -- to which we respond, "so
what?" The type species of the Rhodophyta is probably not Rhodis,
or algal taxonomy is in very deep trouble. Rhodis is the
rhododendron, and not known to be planktonic. Accordingly, we will retain the traditional name for this little
taxon of three genera: Glaucocystis, Cyanophora, and Gloeochaete. Other
genera have been included, at times, based largely on the presence of cyanelles
(primitive chloroplasts). However these appear, on closer inspection, to be
unrelated. In particular, Paulinella is not a glaucophyte, but an
amoeba which apparently acquired its cyanelle by independent primary
endosymbiosis. McFadden
(2001).
Glaucophytes are rather widely-distributed in fresh water, but are never found
in large numbers in any one place. Glaucophytes have a motile stage with two unequal flagellae
of the usual eukaryote type. The flagellae bear two rows of
"hairs," but are morphologically dissimilar to the mastigonemes
of the Alveolata and Chromista. Glaucocystis,
but probably not the other genera, has a cellulose
cell wall. The cell membrane is reinforced by flat vesicles and
microtubules, much like the cortical alveoli found in many alveolates.
Like the red algae, glaucophytes reserve 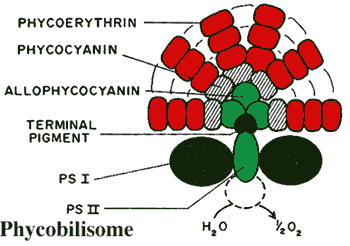 carbohydrates as starch, outside the
chloroplast. The mitochondria are conventional, with flattened cristae.
Glaucophytes undergo open
mitosis and lack centrioles associated with the centrosomes.
carbohydrates as starch, outside the
chloroplast. The mitochondria are conventional, with flattened cristae.
Glaucophytes undergo open
mitosis and lack centrioles associated with the centrosomes.
The glaucophyte chloroplast has the main feature of interest. It retains a number of cyanobacterial features which have been lost in the chloroplasts of red and green algae. In particular, the cyanelles retain a peptidoglycan wall like a eubacterium. The thylakoid membranes, in which photosynthesis takes place, are not stacked, as in green plants, but have a concentric organization. The thylakoids bear clusters of accessory pigments, phycobilisomes, just as cyanobacteria do, with characteristic phycobilin pigments bound to proteins in the same manner as cyanobacteria, as well as other accessory pigments similar to those in bacteria: such as β-carotene, and the carotenoids zeaxanthin and β-cryptoxanthin. The main photosynthetic pigment is always chlorophyll a. Finally, also like cyanobacteria, glaucophyte cyanelles contain carboxysomes, polyhedral structures which stockpile RuBisCO, the enzyme responsible for fixing carbon dioxide. Bhattacharaya et al. (1995); Katz et al. (2004).
Images: Cyanophora from the Phycological Images site of Prof. Isao Inouye, University of Tsukuba. Phycobilisome from the website of Dr. Frank J. Jochem, Florida International University.
ATW050131. Text public domain. No rights reserved.
 There has been a concerted effort in the semi-popular literature to avoid the
word "algae," or to put it into quotation marks, because the algae are
not a monophyletic
group. We have two objections to this practice. First, although the
"algae" are not monophyletic, the red algae, brown algae, etc., are
monophyletic groups. That is, they each include their common ancestor and
all of its descendants. Second, although the "algae" are not
monophyletic, their chloroplasts are monophyletic. That is, all
Cyanobacteria ("blue-green algae") and algal chloroplasts are,
together, all of the descendants of some unique common ancestral
cyanobacterium. Thus, it makes perfectly good phylogenetic sense to speak
of algal chloroplasts; and it is reasonable to call their hosts and prokaryotic
free-living forms algae. The only real problem is that the algae,
thus defined, also include the Embryophyta,
i.e., the land plants. However, we whisk this minor embarrassment under the rug with the observation we are at least consistent with our
discussions elsewhere. From the point of view of phycology, oak trees are
just a peculiar form of green algae. So, other than this explanation, we make no apology for speaking of
Rhodophyta as the red algae.
There has been a concerted effort in the semi-popular literature to avoid the
word "algae," or to put it into quotation marks, because the algae are
not a monophyletic
group. We have two objections to this practice. First, although the
"algae" are not monophyletic, the red algae, brown algae, etc., are
monophyletic groups. That is, they each include their common ancestor and
all of its descendants. Second, although the "algae" are not
monophyletic, their chloroplasts are monophyletic. That is, all
Cyanobacteria ("blue-green algae") and algal chloroplasts are,
together, all of the descendants of some unique common ancestral
cyanobacterium. Thus, it makes perfectly good phylogenetic sense to speak
of algal chloroplasts; and it is reasonable to call their hosts and prokaryotic
free-living forms algae. The only real problem is that the algae,
thus defined, also include the Embryophyta,
i.e., the land plants. However, we whisk this minor embarrassment under the rug with the observation we are at least consistent with our
discussions elsewhere. From the point of view of phycology, oak trees are
just a peculiar form of green algae. So, other than this explanation, we make no apology for speaking of
Rhodophyta as the red algae.
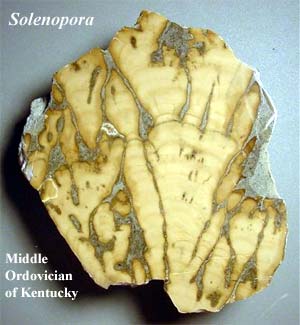
The rhodophytes are an ancient group -- maybe. Some red algae excrete calcium carbonate, so we may have a fossil record to work with. Remains suggestive of red algae have been identified from the Mesoproterozoic. However, these are single-celled forms. Almost all living rhodophytes are multicellular. Supposedly definitive fossils of multicellular red algae are known from the Furongian under the name Solenopora and related forms. We hasten to add that none of these fossil forms appears to be a member of the extant coralline red algae. Worse, a number of recent papers apparently challenge the identification of just about all of the Paleozoic forms. They may be calcareous sponges, cyanobacteria, or stromatolites. On the other hand, the recent identification of Late Proterozoic (or Early Cambrian?) Rhodophyta from superbly preserved phosphate beds in China seems to be a rather sure bet.
 The corallines are still important, arguably the most important, reef
formers today. However, the Rhodophyta are now better known as the source
of nori, and of various gums, gels, glops and goos used as additives to control
the texture and consistency of processed foods, and to prevent the resulting
viscous slurry from separating into its noxious component fluids. If
you've ever wondered what carrageenan
and agar really are, this is it. Red algae were probably the first
multicellular organisms. In fact, few living rhodophytes are
unicells. Over their very long evolutionary lifetime, they seem to have experimented with a
large number of
ways to get cells to stick together. One of the these methods is to
blanket everyone in a paralyzing, gelid mass of sulfonated
sugar polymers -- like institutional tapioca pudding. It works, after
a fashion; but we, as metazoa, are fortunate to have evolved in a different direction.
The corallines are still important, arguably the most important, reef
formers today. However, the Rhodophyta are now better known as the source
of nori, and of various gums, gels, glops and goos used as additives to control
the texture and consistency of processed foods, and to prevent the resulting
viscous slurry from separating into its noxious component fluids. If
you've ever wondered what carrageenan
and agar really are, this is it. Red algae were probably the first
multicellular organisms. In fact, few living rhodophytes are
unicells. Over their very long evolutionary lifetime, they seem to have experimented with a
large number of
ways to get cells to stick together. One of the these methods is to
blanket everyone in a paralyzing, gelid mass of sulfonated
sugar polymers -- like institutional tapioca pudding. It works, after
a fashion; but we, as metazoa, are fortunate to have evolved in a different direction.
Individual cells are also supplied with cell walls, usually of cellulose or xylan (another polysaccharide) fibers. Individual cells are similar to glaucophytes. Sugars are stored as floridean starch -- glycogen, more or less -- which accumulates in the cytoplasm. Rhodophytes, like glaucophytes, lack chlorophyll b but carry a complement of phycobilins arranged in phycobilisomes on unstacked thylakoids. The chloroplasts do not, however, have "cell wall" structures. This system seems to be unusually efficient, as red algae are able to live and photosynthesize at considerable depths, sometimes more than 200m below the surface. Rhodophytes are typically found fixed to substrate in coastal marine environments of almost any kind.
Image: Solenopora from the Kentucky Paleontological Society web site. Solieria from a virtual tour of Chek Jawa.
Links: Divisional Characteristics and Background of Rhodophyta; Introduction to the Rhodophyta; Rhodophyta - The Red Algae; Rhodophyta. The web has many resources on Rhodophyta. But, be warned. For some reason, the proportion of sites offering misinformation of various kinds is unusually high for this taxon.
ATW050204. Text public domain. No rights reserved.
This is the lineage of land plants and green algae. As these have their own section elsewhere, we will not say too much about them here. The Chlorobionta have chlorophyll b and various carotenoid accessory pigments, but lack phycobilins and phycobilisomes. Their cell walls are cellulose, and their storage material is starch, which accumulates inside the chloroplasts. These chloroplasts have stacked thylakoids.
| Page Back | Page Top | Unit Home | Page Next |
