Chordata : Chordata (2)
More urochordates, cephalochordates, and Haikouella
Taxa on This Page
- Ascidiacia
- Cephalochordata
- Thaliacea
DNA and Divergence
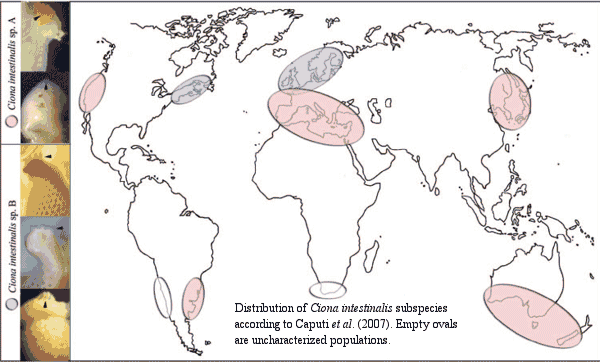 In
2002, the ascideacean Ciona intestinalis became one of the first chordates whose DNA was more or less fully sequenced. The availability of this information is allowing molecular workers to develop new insights into the early evolution of the chordates. However, one of those insights is that C. intestinalis may not be a single species. Caputi et al. 2007) have learned that there are at least two major subspecies of C. intestinalis which cannot cross-breed successfully, although there are few obvious morphological differences. It seems likely that the published DNA sequence is entirely from Caputi's "subspecies A", since this variant is the sole type in both California and Japan, the locations where the sequencing was performed. Other subspecies, or perhaps even intermediate forms may remain to be characterized.
In
2002, the ascideacean Ciona intestinalis became one of the first chordates whose DNA was more or less fully sequenced. The availability of this information is allowing molecular workers to develop new insights into the early evolution of the chordates. However, one of those insights is that C. intestinalis may not be a single species. Caputi et al. 2007) have learned that there are at least two major subspecies of C. intestinalis which cannot cross-breed successfully, although there are few obvious morphological differences. It seems likely that the published DNA sequence is entirely from Caputi's "subspecies A", since this variant is the sole type in both California and Japan, the locations where the sequencing was performed. Other subspecies, or perhaps even intermediate forms may remain to be characterized.
Recently, several species of the fruit-fly Drosophila, have been fully sequenced in tandem. Using these sequences, it will be possible to look closely at the types of genetic changes which are involved in the initial divergence and radiation of new species in that model system. The existence of cryptic speciation in Ciona will be a valuable check on these results. Earlier work in plants (Arabidopsis) and fungi (Neurospora) suggests that the types of genetic changes involved in speciation may not be universal across phylospace. That is, at the genetic level, evolution may or may not) work in quite different ways for widely separate types of organisms.
One early lesson from these comparative studies seems to be that gene rearrangement is at least as important as gene mutation in the formation of new species. The preliminary findings of Caputi et al. are consistent with this pattern. However, it remains to be seen whether gene rearrangements simply cause gross errors during chromosomal exchange in meiosis, or whether they also implicate more subtle rules relating to gene regulation. ATW080123.
Descriptions
 Ascidiacea:
Ciona, Distaplia, Diplosoma. Classic urochordates with "tadpole" larvae and sessile adult.
Ascidiacea:
Ciona, Distaplia, Diplosoma. Classic urochordates with "tadpole" larvae and sessile adult.
Adult has incurrent and excurrent siphons and uses branchial filter feeding; motile larva has notochord, dorsal nerve chord, segmented muscles, and tail; solitary, social & compound forms.
Thaliacea: Salps. Free-living forms morphologically like Ascidiacean adults
Free-living forms morphologically like Ascidiacean adults.
Links: Thaliacea: brief checklist; Dive Log New Zealand : Phylum Chordata. APW 040314.
Fusiform, <5cm. Brain segmented & perhaps functionally homologous to vertebrate brain; segmented muscle (myomeres) pull alternate sides, stiffened by notochord. Pharynx & gills used only for filter feeding. Circulation limited, but ventral pump & major dorsal artery. No hemoglobin; oxygen carried in solution; no kidney, but similar excretory cells. No connection between segmented & unsegmented nerves. Profound left-right asymmetry in development. Semi-sedentary filter feeder, trapping detritus in mucus; water moved by cilia – mouth and gut have little or no musculature. No paired fins, thus little or no control in 3 dimensions. Neural crest cells absent.
Links: Chordata introduction - Subphylum Cephalochordata; Characteristics; Chordata; Berkeley. APW
040314.