
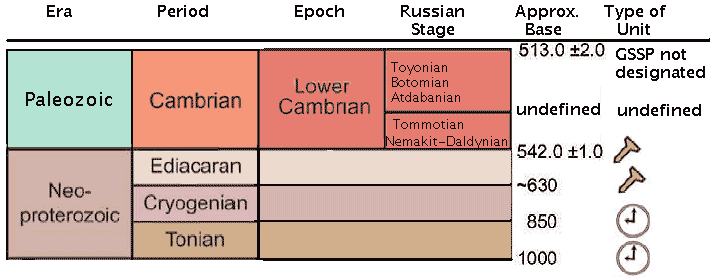
| Ediacaran Period | ||
| Neoproterozoic | Ediacaran - 1 |
| Cryogenian | Mesoproterozoic | Neoproterozoic | ||
| Cambrian | Paleozoic | Timescale |
This page describes the stratigraphy and fossil record of the Ediacaran (f/k/a Vendian) Period, including a brief introduction to the Ediacaran fauna. Some famous Ediacaran Lagerstätten are briefly discussed, followed by a sketched outline of some of the major evolutionary events.
The Ediacaran Period and System were first proposed by Sokolov 1952, from drill core sequences on the Siberian Platform. Sokolov & Fedonkin (1984). Although the Ediacaran was not embraced quickly, it is now recognised by the Subcommission on Precambrian Stratigraphy and has now come into almost universal usage -- although frequently still called the Vendian.
As with the better-known Cambrian Period, the absolute age constraints on the Ediacaran interval have ebbed and flowed over the past few years; the best current best guess is from 630 to 542 million years (Ma) ago.
The Precambrian era was a period in earth history before the evolution of hard-bodied and complex organisms. Throughout the extent of both periods, dominant Precambrian and Ediacaran organisms were simple, entirely marine, and for the most part soft-bodied: hard-bodied organisms did not occur until nearly the beginning of the Cambrian Period when the so-called "small shelly faunas" appeared.
The Ediacaran Period and System were first proposed by Sokolov 1952), from drill core sequences on the Siberian Platform. It was officially adopted by the ICS in 2004. The base of the Ediacaran (the Global Standard Stratotype Section and Point, or GSSP) is defined by reference to the base of the Marinoan cap carbonate (Nuccaleena Formation), immediately above the Elatina diamictite in the Enorama Creek section, Flinders Ranges, South Australia. See discussion at The Ediacaran Period- A New Addition to the Geologic Time Scale.
While we follow the ICS designation, we agree with the dissenting opinion of the Russian members. This may not have been the best pick for a number of reasons:
1) The age of the GSSP is very poorly constrained.
2) At least one widespread "Ice Age" post-dates the GSSP.
3) There are no signs of metazoan life anywhere near the GSSP, much less any Ediacaran assemblage.
4) the name is confusing, since it suggests an Ediacaran assemblage.
Since 1947, when H.E. Wheeler initiated debate with the suggestion that the Precambrian-Cambrian boundary should be based upon the first appearance of trilobites, much has ensued. Progress has largely been facilitated by the International Geological Congress (IGC) and the establishment in 1960 of a Subcommission on Cambrian Stratigraphy. The classical idea of placing the boundary at an unconformity has been displaced by the search for monofacial, continuous deposition sequences across the boundary, with the view to selecting a stratotype.
The search itself produced a wealth of data from around the world – including the Palaeotethyan Belt, Siberian Platform, and England – eventually focusing upon south-east Newfoundland. In 1991 the International Subcommission on Cambrian Stratigraphy through its Working Group on the Precambrian-Cambrian Boundary) made the official decision to draw the base on the Cambrian at the first appearance date (FAD) of Trichophycus pedum (fig. 1) in the reference section at Fortune Head.
Fig. 1: The horizontal burrow trace fossil, Trichophycus (formerly Phycodes) pedum defines the lower boundary of the Cambrian in the reference section at Fortune Head, southeastern Newfoundland. [Image courtesy of Dr. Gerd Geyer, Institut für Paläontologie, Bayerische Julius-Maximilians-Universität, Würzburg, Germany.]
 The name "Ediacaran," in its geochronologic sense, used to mean an upper subunit of the Vendian, approximately 565 to 543 Ma (Bowring & Erwin 1998), with a stratotype in South Australia. This was supposed to take the age of the Ediacaran fauna, while the Varangian sub-period took in the age of the Varangian glaciation -- presumably the first half of the Vendian.
The name "Ediacaran," in its geochronologic sense, used to mean an upper subunit of the Vendian, approximately 565 to 543 Ma (Bowring & Erwin 1998), with a stratotype in South Australia. This was supposed to take the age of the Ediacaran fauna, while the Varangian sub-period took in the age of the Varangian glaciation -- presumably the first half of the Vendian.
Unfortunately, both nomenclature and dating have changed considerably:
The Ediacaran is still the age of the Ediacaran fauna, sort of. We can conceive of the Ediacaran as a period during which the Ediacaran fauna presumably evolved and in which conditions were generally right for it to live, even if we have yet to establish its presence with certainty. The earliest signs of metazoan life at Chengjiang almost reach the 600 My mark, and the earliest Twitya trace fossils push 610 My. The GSSP lies somewhere between 600 and 635 Ma, with the most likely date being 630 Ma. Thus the correspondence between the GSSP and metazoan life is not all that unreasonable. The Ediacaran-Cambrian boundary (i.e. the base of the Cambrian, is still dated at 542 Ma -- about the same as it has been for the last decade.
The Cryogenian is a chronostratigraphic unit. That is, its base is defined in terms of years. It cannot move, and it has no GSSP. Thus its base is fixed at 850 Ma and its end is defined as the beginning of the Ediacaran, or about 630 Ma.
The "Varangian" is not a recognized unit. Fortunately, the age of the Varanger glaciation has also been re-dated -- at 800 to 630 Ma. Thus, the Cryogenian covers essentially the same turf. Despite the sanction of the ICS, the term "Varanginian" is used far more often than "Cryogenian." The broad (but short-lived) Ice Age which occurred about 600 or 580 My is a distinct event. The tendency in the recent literature has been to call it the Marinoan and to distinguish it from the world-wide Varanger (or Varanginian) Ice Age which ended about 630 My.
The "Vendian" no longer exists as a stratigraphic unit. This is a good thing. Personally, we liked the name. It was easy for even Texans to pronounce. It honored the important Russian contributions to Pre-Cambrian stratigraphy. It was more widely used than "Ediacaran," and less easily misspelled. However, it was always hopelessly ambiguous. We suspect the Russians will have their day when the Early and Middle Cambrian ages are finally agreed on.
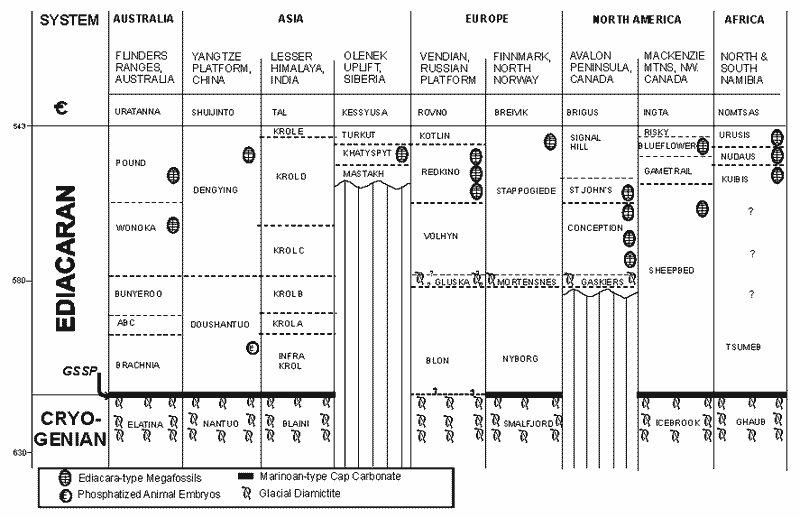
The following more detailed look of the stratigraphy of the Ediacaran comes from the Report of the Terminal Proterozoic Subcommission of the International Commission on Stratigraphy, Knoll et al. (2000):
Ask any Texan, even your normally urbane and cosmopolitan host, about the Middle East. We will be happy to share our knowledge of the fabled realms of Eye-Rak, Gutter (Qatar), Sow-Dee, Jawd'n (1 syllable), and Iz-rul.
The Precambrian supercontinent usually known as Rodinia (or, rarely, as Proto-Pangea or Ur-Pangea) formed ~1,000 Ma from the amalgamation of three or four pre-existing continents, in an event known as the Grenville Orogeny. Perhaps beginning ~700 Ma, but protracted over many millions of years, Rodinia began breaking up into three major blocks: West Gondwana, East Gondwana, and Laurasia. Subsequently - perhaps ~540 Ma - West and East Gondwana merged in the mountain-building event known as the Pan-African Orogeny. (After Rogers 1996.)
605 to 585 Ma: The Varanger-Marinoan Ice Ages
It has been suggested that the Varanger-Marinoan ice ages, which lasted from approximately 605 to 585 Ma (Martin et al. 2000), were "snowball" events in which glaciation extended to very low latitudes; possibly right to the equator. It may have been a time of widespread extinction, a contention based mainly on carbon isotopic profiles, which display large negative excursions. Anyone who thinks they understand the "snowball" phenomenon should consult the defense of our strongly held position of total confusion on this subject.
Post-Glacial Ediacaran
The post-glacial Ediacaran was warm to hot and relatively arid at low and most middle latitudes. Even at high latitudes in the south, there is evidence of a warm, humid climate. The latest Ediacaran and earliest Cambrian were marked by a return to colder conditions, with some glaciation at high latitudes. (mod. ATW060108)
Trace fossils
Body fossils typically of cnidarian grade dating from as early as 600 or 610 Ma - e.g. the Twitya fossils are simple cup-shaped animals, possibly similar to the sea anemones of today.
Mineralised skeletons of uncertain affinity - the 'small shelly fauna' - appear just before the beginning of the Cambrian, ~550 Ma, increasing in numbers and diversity towards the Tommotian. The most common skeletal materials are calcium carbonate (aragonite or calcite) and varieties of calcium phosphate. Many of the latter may originally have been carbonates, phosphatized during preservation. The oldest of these to occur abundantly are Cloudina and the allied genera comprising the family Cloudinidae: small, conical fossils made of calcium carbonate, first (?) appearing in the Ediacaran Stirling Quartzite of California (Langille, 1974) and persisting into the Cambrian. Anabarites and Cambrotubulus are other Ediacaran SSF taxa, known from Siberia and Mongolia.
Simple disc-like impressions, interpreted as cnidarian-grade body fossils, have been reported from the inter-tillite beds of the 610 to 600 Ma Twitya Formation, in the Bluefish Creek area of the McKenzie Mountains, north-western Canada. Hofmann et al. (1990). The Twitya Formation is a ~800 m thick succession of siliciclastic turbidites within the Windermere Supergroup. Fossils occur in the interval 170 to 200 m below the top of the formation.
The age of the assemblage (approximately 610 Ma, Martin et al. 2000: fig. 1), indicates deposition during the early part of the Varanger-Marinoan ice age. This placement is supported by overlying deep-water diamictite containing dropstones, rare striated clasts, and relict ice-cemented boulders.
The Twitya impressions are preserved as "very simple, convex discoidal and annular reliefs on the lower surfaces of thin sandstone beds, and as counterparts on the tops of the underlying shale beds." Hofmann et al. (1990: 1199). This mode of preservation is quite characteristic of the Ediacaran fauna, which typically occurs in somewhat younger rocks. Inorganic sedimentary structures such as gas-evasion marks, load and dewatering structures, are common in the Twitya Formation. However, systematic differences between the presumed fossils and obviously inorganic structures, as well as a high degree of morphological consistency exhibited by the hundred or more specimens, argue strongly for a biogenic origin.
Hofmann et al. provisionally assigned the fossils to three taxa: Nimbia occlusa, Vendella?, and Irridinitus? The last two of these, however, are both probably junior synonyms of Aspidella terranovica (refer Gehling et al., 2000: 448) so probably only two taxa are represented. Their occurrence at the bottom of turbidite beds suggests a sessile habit. They are most widely interpreted as the basal impressions of cnidarians – either simple pedestal impressions or possibly the remains of a sand 'ballast' retained within the organisms during life – or at least as metazoans of cnidarian grade. "Interpretation as colonial aggregates of prokaryotes e.g. Nostoc-like balls) is possible but is difficult to reconcile with the morphology and relatively high relief of the remains, their occurrence at the bottom of turbidite beds, and the lack of a carbonaceous film outlining them, particularly in view of the of the fact that carbonaceous compressions are present in the formation." Hofmann et al. 1990: 1202).
The principal significance of this occurrence of cnidarian-grade metazoans is their stratigraphic position below ?Varangian glaciomarine tillites (Aitken 1988, 1989). This is the only Ediacaran-like assemblage found below Varanger glacial deposits anywhere, and provides a useful test for those models positing metazoan evolution to have been arrested during one or more of the Neoproterozoic glaciations. See, e.g., Runnegar (2000); Peterson & Davidson (2000).
In addition to the putative cnidarian impressions, the Twitya Formation contains the carbonaceous film taxa, Morania and Beltina, and also some poorly preserved filamentous microfossils and leiosphaerids.
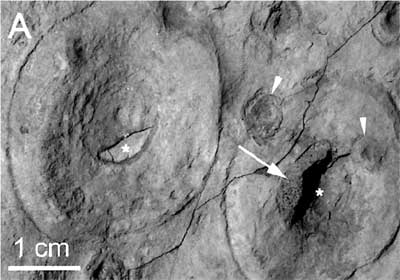
Several examples of Aspidella in the Fermeuse Fm. of Newfoundland, Canada, from Peterson et al. (2003). These authors make a strong argument, based on mode of life and size distributions, that Aspidella was a fungal growth form. They expressly disclaim any assertion that all of the Ediacaran forms were fungi. However, one quite plausibly imagine a Late Neoproterozoic in which sponges, early cnidarians, and fungi contested for ecological dominance.

Possible Nimbia specimen from the Ediacaran of Norway. Farmer et al. (1992). Note that both Nimbia and Aspidella may be holdfast marks left by some organism or colony with a stem and frond-like extensions, as in Spriggina.
This requires a little explanation. Refer to the figure in Knoll et al. (2000). As we hinted, there is a separate glaciation event which shows up in some exposures and dates to between 600 and 580 My.Some of the Twitya trace fossils apparantly occur below the tillite associated with this Ice Age, which some call the Marinoan. So far as we are aware, no one claims to have found metazoans from a level below the Ediacaran GSSP.
An impoverished but characteristic Ediacaran assemblage occurs in the upper beds of the Drook Formation, south-eastern Newfoundland, 1500 m stratigraphically below the well-known Mistaken Point fossils; these are the oldest of the large, architecturally complex fossils found so far (Narbonne & Gehling, 2003). The published age constraints on these fossils are from 595 Ma (Varangian glacial diamictites of the Gaskiers Formation) to 565 Ma (well-dated Ediacaran fossils at Mistaken Point occurring 1.5 km stratigraphically higher). Unpublished data noted in Walker (2003: 220) indicates an age of 575 Ma.
Current-aligned fronds attributable to the cosmopolitan Ediacaran, Charnia masoni, and those of a large (up to nearly 2 m in length) new species, Charnia wardi, occur on the shaly tops of turbidite beds under volcanic ashes. Their position above the glacial marine rocks of the Gaskiers Formation (595 Ma) provides our earliest window on life following the Varanger ice age.

Holotype of Charnia wardi from the Ediacaran of the Drook Formation, Newfoundland, Canada. From Narbonne & Gehling (2003).
Soft-tissue fossils preserving cellular structures, notably including the earliest record of sponges (see below), occur in the Doushantuo Formation phosphates, exposed near Weng'an in Guizhou (south central China), providing evidence of a diverse biota.
Age constraints on the Doushantuo Formation are rather weak. Chemostratigraphic profiles suggest that Doushantuo fossils predate the last strongly positive carbon isotope excursion of the Proterozoic, dated as 549 ± 1 Ma in Namibia (Grotzinger et al. 1995). Similarly, Doushantuo microfossils provide biostratigraphic evidence that this formation predates the 555 ± 3 Ma sandstones of the Redkino Series, northern Russia, which contain diverse Ediacaran body and trace fossils. Bio- and chemostratigraphic correlations further suggest that Doushantuo fossils are older than diverse Ediacaran assemblages found in Australia, Ukraine, and northern Siberia. However, in the absence of direct radiometric constraints, it is uncertain whether Doushantuo fossils predate frondose Ediacaran remains from Newfoundland, dated at 565 ± 3 Ma, although even the age of 570 Ma for the Doushantuo fossils proposed by Martin et al. (2000) (much less the c. 600 Ma found by Knoll et al., 2000) places them some 5 Ma earlier. Knoll 2003, p. 141, suggests a range of 600 to 590 Ma. Whichever is correct, the deposit seems certain to be post-Varangian.
The reported biota now includes probable algae, cnidarians and bilaterians – the last two largely known from fossil embryos. Unfortunately, diagenetic effects are sometimes difficult to distinguish from genuine biological structures, and much of the evidence from this source, though widely accepted, remains equivocal.
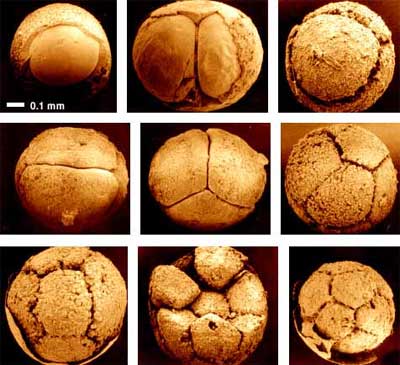
Embryos from the Doushantuo Formation. Image from the website of Prof. Shuhai Xiao, Virginia Tech Univ.
The oldest of the diverse Ediacaran assemblages yet described is that from Mistaken Point, eastern Newfoundland, where fossils are spectacularly preserved on large bedding surfaces along the sea-cliffs of the Avalon Peninsula. Zircons from interbedded ash have been dated at 565 ± 3 Ma (Benus 1988).
The Mistaken Point assemblage contains a few cosmopolitan taxa such as Charnia and Aspidella, but most are either endemic or shared only with the Charnwood Forest locality in central England.
The The two most abundant and diverse Ediacaran trace and body fossil assemblages are those from the White Sea coast of Russia and from the Flinders Ranges in South Australia, which together account for 60% of the well-described Ediacaran taxa.
"Many exposures in the White Sea region contain known Ediacaran biotas; however, the best fossil occurrences are found along the shoreline cliffs at Zimnie Gory. These unmetamorphosed and nondeformed (except for present-day cliff-face slumping) siliciclastic rocks belong to the uppermost Ust-Pinega Formation and form the northern flank of the Mezen Basin along the southeast flank of the Baltic Shield." Martin et al. (2000: 842). Zircons from a volcanic ash in the lower part of the sequence preserved between Medvezhiy and Yeloviy Creeks (id.) yielded a date of 555.3 ± 3 Ma, the minimum age for the "oldest definitive triploblastic bilaterian, Kimberella, and the oldest well-developed trace fossils; and it documents that spectacularly diverse and preserved Ediacaran fossils formed more than 12 million years before the base of the Cambrian." Id. at 843.
"The fossil Kimberella quadrata was originally described from late Precambrian rocks of southern Australia. Reconstructed as a jellyfish, it was later assigned to the cubozoans ('box jellies'), and has been cited as a clear instance of an extant animal lineage present before the Cambrian. Until recently, Kimberella was known only from Australia, with the exception of some questionable north Indian specimens. We now have over thirty-five specimens of this fossil from the Winter Coast of the White Sea in northern Russia. Our study of the new material does not support a cnidarian affinity. We reconstruct Kimberella as a bilaterally symmetrical, benthic animal with a non-mineralized, univalved shell, resembling a mollusc in many respects. This is important evidence for the existence of large triploblastic metazoans in the Precambrian and indicates that the origin of the higher groups of protostomes lies well back in the Precambrian." Fedonkin & Waggoner (1997).

Kimberella from the Winter Coast of the White Sea, Russia. Image from Erwin & Davidson (2002). These authors note the presence of certain "adjacent parallel lines [which] are trace fossils associated with Kimberella, and are believed to represent infilled feeding scratches through a microbial mat. The presence of these feeding traces suggests that Kimberella possessed feeding structure similar to the molluscan radula." Id.
Further Reading
The Precambrian-Cambrian Boundary – Cowie, J.W. and Brasier, M.D. (eds.)
Related Pages
Other Web Sites
page uploaded 020515
unless otherwise indicated, text © Chris Clowes 2002
last revised ATW060108, edited RFVS111126
checked ATW031227