Pteraspidomorphi: Amphiaspidida
Ctenaspis
Taxa on This Page
- Amphiaspidida X
- Ctenaspis X
Amphiaspidida
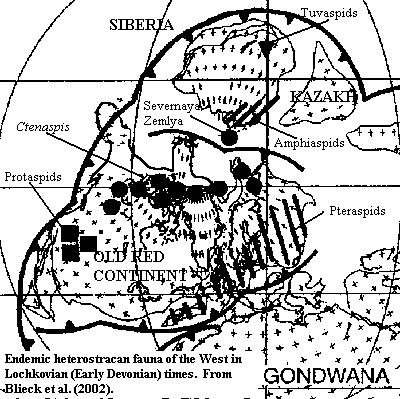 In the Pridoli and during the Lochkovian Age (the earliest age of the Early Devonian) vertebrates were first becoming major elements of the world fauna. The process was a remarkably slow one. In fact, for perhaps the last time in vertebrate history, the rate limiting factor in global faunal change was dispersal, rather than evolution. By way of contrast, consider the tetrapod radiation which began at the end of the Devonian. We really have no idea where our ancestors first walked on land because, as soon as that lineage became successful, its members are found all over the world. Tetrapods radiated in the blink of a geological eye. Undoubtedly, this "blink" took a few hundred thousand years, but, from our standpoint that is virtually instantaneous. Marine vertebrates, in particular, are very mobile. If they are found anywhere, they tend to be found everywhere there is any chance of dispersal.
In the Pridoli and during the Lochkovian Age (the earliest age of the Early Devonian) vertebrates were first becoming major elements of the world fauna. The process was a remarkably slow one. In fact, for perhaps the last time in vertebrate history, the rate limiting factor in global faunal change was dispersal, rather than evolution. By way of contrast, consider the tetrapod radiation which began at the end of the Devonian. We really have no idea where our ancestors first walked on land because, as soon as that lineage became successful, its members are found all over the world. Tetrapods radiated in the blink of a geological eye. Undoubtedly, this "blink" took a few hundred thousand years, but, from our standpoint that is virtually instantaneous. Marine vertebrates, in particular, are very mobile. If they are found anywhere, they tend to be found everywhere there is any chance of dispersal.
The case was different at the beginning of the Devonian. Most vertebrate faunas were endemic, that is, local. Protaspidoids dominated in western Laurentia at the western end of the "Old Red Continent" of Laurasia. Pteraspidids inhabited Baltica in the East, and a mixture of Ctenaspis, Gigantaspis and various osteostracans held the equatorial zone around what is now arctic Canada. Blieck et al. (2002). Each of these lineages radiated separately, with genus succeeding genus for the 10-20 million years of this era. But the process of dispersal was so slow that the rate of faunal replacement between different groups was much slower than the process of evolution within them.
Of all the endemic groups, the Amphiaspidida were one of the most stubbornly provincial. They are usually described as inhabiting "Siberia." But that is a gross exaggeration. Amphiaspids are found only in the "Kara-Tajmyr" block, what is now a relatively small strip across the central part of extreme northern Russia. At least one other fauna inhabited the Siberian Platform, the completely unrelated Tuvaspid fauna of Tuva.
The amphiaspids are often described as "bizarre," which is unkind, but accurate. With rare exceptions, the head shield is entirely fused into a seamless coat of bone, typically bearing a sort of brim laterally which was often incised into a series of jagged "teeth." As in all heterostracans, the eyes were small and without sclerotic rings. But some hibernaspidoid forms were completely eyeless and others bore tiny eye spots at the end of a long oral tube, like a built-in straw. In hibernaspidoid and amphiaspidoid species with more or less "normal" eyes, the orbit is partially rimmed by a second, bean-shaped opening of unknown function. It is widely speculated that this opening had an inhalant function. What's particularly odd is that the same sort of structure is found in the very primitive, Ordovician fish, Sacabambaspis, but is otherwise unknown. There is no real possibility of a direct relationship. Amphiaspids are a late and short-lived breed. Except for the stem-amphiaspid Ctenaspis, they appear only in the latter part of the Lochkovian and are gone by the Middle Devonian.
Amphiaspids are typically depicted as bottom-feeding detritivores. Yet, even more than most heterostracans, they tend to be flat on top with convex, even inflated, ventral surfaces. That sort of shape is hard to reconcile with the usual benthic lifestyle. Even if partly buried in sediment, any current would tend to dig these boat-shaped bodies out and lift them off the bottom. Ironically, this is particularly true of some of the blind, tube-mouthed Hibernaspidoidei, such as Eglonaspis. One amphiaspid, Siberiaspis, appears to have had two lateral line systems: a primitive superficial pit line network, and a deeper set of channels. Halstead 1973) (reviewing work of Novitskaya). This tends to reinforce a point made earlier, in connection with the Corvaspididae. The various layers of the heterostracan integument may, to a significant degree, have had independent evolutionary histories consistent with their distinct developmental origins. ATW030513.
 Amphiaspidida: (same as Amphiaspida and Amphiaspidae of older sources, more or less the amphiaspidiformes of other sources)
Amphiaspidida: (same as Amphiaspida and Amphiaspidae of older sources, more or less the amphiaspidiformes of other sources)
Early Devonian (Lochkovian to Emsian) of Siberia. Survival to Pragian of Middle Devonian per [J96]. Reports of amphiaspids in Arctic Canada and Spitsbergen are based on the occurrence of Boothiaspis, which is now believed to be a cyathaspid [B+02]. However, this range is still correct if Ctenaspis is an amphiaspid [BJ99]. All occurrences are from warm, marine waters. [BJ99].
Characters: up to 40 cm, but typically 10-18 cm [L95]; wide, rounded carapace [N86] [L95] (contra [N86]: may be straight anterior border); armor wide & deep [S64]; armor about as wide as long or slightly longer than wide [S64]; some with elongate, tube-like rostrum bearing orbits as well as mouth [N86] [J96] [BJ99]; mouth ventral or terminal, sometimes at the end of an oral tube [S64] [N86]; no supraoral plate [S64]; head is "much flattened" [J96]; loss or reduction of eyes [N86] [L95] [BJ99]; orbits (if not absent or on rostral tube) small, dorsal and usually far anterior [S64]; orbit lined externally with bean-shaped secondary opening which may be functional spiracle [J96]; gill opening usual dorsal or lateral, and rarely on a separate branchial plate (Putoranaspis) [H73] [N86]; dorsal surface generally flat [H73]; dorsal surface usually bears a crest or spike [N86]; complete fusion of carapace [N86] [L95] [J96] (contra [H73]: some forms have separate dorsal & ventral plates); posterior ventral area sometimes not fused [N86]; in some cases, distinct ornament borders are present between the dorsal, ventral and branchial elements -- and rarely on borders of the suborbital plates (Aphataspis, Gerronaspis) [N86]; lateral edges of armor are smooth or with jagged "teeth" [N86]; jagged processes may also be present on the posterior margin [S64]; along lateral edges armor forms a flattened rim [N86]; posterior margin, as a rule, has a median ledge [N86]; ventral surface convex [H73]; sensory line is system propagated by furrows and channels, including 'V'-shaped supraorbital channels, lateral channels, longitudinal lines, and cross-commisures [N86]; ventral cross-commisures unique to this taxon [S64]; some forms appear to have had an internal, as well as external sensory line [H73]; ornament of dentine, ridges and mounds with smooth or jagged edges [N86]; ornament of stellate tubercles (= grebeshki?) [S64]; mounds alternate with ridges or entirely cover small areas of the armor [N86]; ridges are generally oriented longitudinally, and sometimes (on the periphery of the armor) -- radially [N86]; some forms are tesserated [N86]; in an average part of an armor, there is usually one layer of prominently prismatic chambers [N86].
Note: [1] some specimens known with healed bites, presumably from gnathostomes. [L95].
Links: Heterostraci; 420 Text; MsoDockBottom; Biology 356; Upper Silurian and Devonian heterostracan pteraspidomorphs ( ...;
References: Blieck & Janvier (1999) [BJ99]; Blieck et al. (2002) [B+02]; Halstead 1973) [H73]; Janvier 1996) [J96]; Long (1995) [L95]; Novitskaya (1986) [N86]; Stensiö (1964) [S64]. APW040316.
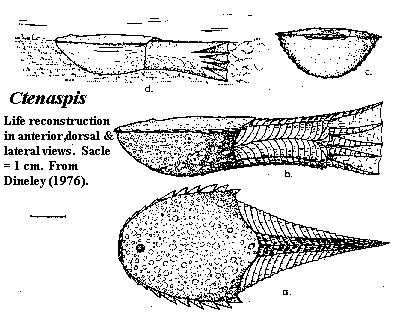 Ctenaspis: C. dentata Kiaer 1930; C. cancellata Kiaer, 1930; C. kiaeri Zych, 1931; C. obruchevi Dineley, 1976; C. ruselli Dineley, 1976; C. ornata Dineley, 1976.
Ctenaspis: C. dentata Kiaer 1930; C. cancellata Kiaer, 1930; C. kiaeri Zych, 1931; C. obruchevi Dineley, 1976; C. ruselli Dineley, 1976; C. ornata Dineley, 1976.
Early Devonian (Lochkovian
- Pragian) of North America (Arctic Canada), Lochkovian of Spitzbergen, Siberia (Severnaya Zemlya), England and Ukraine (Podoli) [D76] [N86] [BJ99]
Phylogeny: Amphiaspidida: (Amphiaspidoidei + (Hibernaspidoidei + Siberiaspidoidei)) + *.
Characters: <10 cm?; adorbital opening (praespiraculum) absent [J96]; only two (dorsal & ventral) plates [D76]; dorsal plates fused with branchials [N86]; plates notably thin [D76]; dorsal plate with jagged lateral margins [N86]; dorsal plate with well-developed lateral brim [D76]; armor rounded and slightly lengthened [N86]; shield short & broad (l/w = 0.7-0.9) [D76]; dorsal shield flat [D76]; short, separate supraoral shelf present with fin anteroposterior striations [D76]; preorbital process well-developed [N86] [1]; orbits small and located relatively far anteriorly [D76]; pineal macula well-developed [D76] [N86]; branchial opening located at posterior edge of armor, between dorsal & ventral plates [D76] [N86]; posterior edge of dorsal shield with triangular ledge ornamented with a network of ridges [N86]; ventral shield strongly arched except marginally [D76]; epitega & scale-like units absent [S64] [N86] [SW90]; dentine ridges absent [N86] [SW90]; ornament of stellate tubercles [J96]; ornament of aspidine tubercles (not dentine [D76] [3]) and local networks of ridges [N86]; ornament notably coarse [D76]; ornament includes either flat, wide tubercles with a pointed end posteriorly, or a network of delicate ridges [S64]; sensory canals tend to branch [N86]; some  elongated scales rows present on body [SW90]; tail diphycercal [J96]; tail posterior margin straight or slightly concave [SW90]; dorsal & ventral ridge scales terminating in points [SW90]; up to 6 large "rays" each consisting of a single row of posteriorly tapered scales, supporting fin web [D76] [SW90]; probably no separate muscular control of fin rays [J81]; .
elongated scales rows present on body [SW90]; tail diphycercal [J96]; tail posterior margin straight or slightly concave [SW90]; dorsal & ventral ridge scales terminating in points [SW90]; up to 6 large "rays" each consisting of a single row of posteriorly tapered scales, supporting fin web [D76] [SW90]; probably no separate muscular control of fin rays [J81]; .
Note: [1] not an elongate rostrum as in most amphiaspids, but a broad, rounded anterior extension of the dorsal plate, more in the manner of pteraspidiforms. [2] Dineley [D76] reconstructs Ctenaspis as benthic, but a capable swimmer apparently because the tail, being flexible from top to bottom, might have been used for attitude control. [3] Stensiö [S64] explains: the dentine layer was reduced in Ctenaspis so that the ornament was largely raw bone. Presumably this helps account for the unusually thin plates. Stensiö also analyzes the ornament at great length in terms of primitive cyclomorial units. Following Janvier [J96], I have omitted almost all such discussion as probably pointless.
Links: Upper Silurian and Devonian heterostracan pteraspidomorphs ( ....
References: Blieck & Janvier (1999) [BJ99]; Dineley 1976) [D76]; Janvier (1981) [J81]; Janvier 1996) [J96]; Novitskaya (1986) [N86]; Soehn & Wilson (1990) [SW90]; Stensiö 1964) [S64]. ATW030509.