 Aspidine or, sometimes, "aspidin")
Aspidine or, sometimes, "aspidin")| Pteraspidomorphi | ||
| The Vertebrates | Psammosteida (1) |
| Vertebrates Home | Vertebrate | Vertebrate |
Abbreviated Dendrogram
Vertebrata
├─Conodonta
└─┬─Thelodonti
│
└─Pteraspidomorphi
├─Arandaspida
└─Heterostracomorphi
├─Astraspidae
└─Heterostraci
├─Cyathaspidiformes
│ ├─Cyathaspidida
│ └─Amphiaspidida
└─Pteraspidiformes
├─Pteraspididae
└─Psammosteida
├─Drepanaspis
└─Psammosteidae |
Contents
Overview |
"The liveliness of controversies engendered by these animals does not appear to have diminished one iota during the last 150 years. It must be a cause of some surprise that these apparently inoffensive fish should be capable of arousing such passions." Halstead 1973)
In these Notes, we seldom address questions such as "why do we care?" Anyone who reads these words has already manifested a disturbing degree of interest in the arcana of evolutionary biology, as well as a willingness to seek it out in unlikely places. One is not likely, for example, to merely stumble on psammosteid histology while searching for celebrity images in varying stages of undress or reproductive receptivity. Second, there are many answers to the question. Some conflict. Some touch on matters which set off crackling visceral reflex arcs, hard-wired deep within the political brainstem of the academic mind. Nevertheless, histology is one of those subjects which, even by the notably lax standards of paleontology, is considered dull, obscure and old fashioned. Thus its presence here requires some sort of justification.
Histology is a word with a phylogeny sometimes more interesting than the science itself. The Greek root seems to be related to hoist, with precisely that meaning. The sequence then became: to step (a mast) or raise up > a mast or pole > anything raised up > a (vertical) loom > cloth made on a loom > any web or (living) tissue. With the development of microscopy and artificial dyes, histology became the molecular biology of the Nineteenth and early Twentieth Centuries. There is still a vast arcana of strange, evocative stain terminology: the "Gomori-Takematsu Procedure," "Congo Red," and "Weigert's Iron Haematoxylin." Much of this sounds as if it belonged on an alchemist's parchment or perhaps in some New Age yoga manual. The work itself was highly empirical, "let's add a little of this stuff and see what happens," bench-top chemistry, subject to many artifacts and sometimes requiring an extraordinary amount of individual skill, experience and good luck.
With the advent of more contemporary biochemistry and molecular biology, bizarre acronyms ("cAMP-activated Ca-dependent GTPase") and genetic silliness ("Sonic Hedgehog") displaced most of this odd, but dignified, old vocabulary, as well as the odd, but dignified, old professors who had used it. Much was gained by this transition. However, something was lost as well. Few scientists currently work at the tissue level of organization. We have made enormous progress in understanding the molecular basis of life, and are even beginning to assemble this knowledge to attack the cellular level. At the other end of the spectrum, organismal biology and ecology are making a modest comeback. At least by comparison, physiology (organ-level) and histology tissue-level) studies are presently languishing. Unfortunately, in some cases, tissues are almost all we have. The psammosteids are one of those cases. Most psammosteids consist of pieces of dermal plate, more or less crushed into fragments. Absent some incredible new Devonian lagerstät, that is all we will ever have [1]. If we are to understand this group, we must look seriously at their histology.
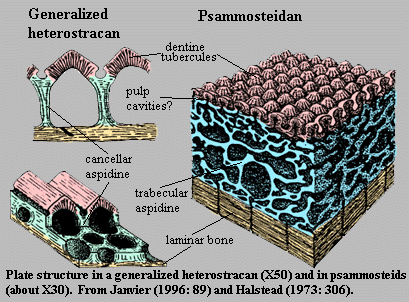
Aspidine or, sometimes, "aspidin")The plates of psammosteids, like those of other pteraspidomorphs, are made up of aspidine. The difficulty is that no one can define this term. Histology, particularly as applied to fossil material, does not distinguish adequately between bricks and brick walls. That is, the literature does not draw a bright line between the chemical composition of aspidine and its structure. Janvier 1996) suggests that aspidine is simply a term for acellular bone. This is useful but, ultimately, only adds a layer of developmental debate to the already existing confusion between structure and composition. Furthermore, Halstead (Tarlo (1964)) argues that psammosteidan plates are not acellular [2].
Since we are dealing with materials from fishes that have been dead, and buried under tons of rock, for some four hundred million years, we cannot assume that their chemistry has been unaffected (even if we knew what it is today). We certainly cannot observe the developmental biology of psammosteidan integument. We can, however, observe that their remains are structurally distinctive, although plainly related to other pteraspidomorph plates. Accordingly, a structural definition seems the most workable.
On a structural level, it is plain that the psammosteidan integument differs from aspidine as it is known from heterostracans (i.e. Devonian pteraspidomorphs). As Halstead discusses at great length, in some ways it more closely resembles the exoskeleton of the very earliest pteraspidomorphs, such as Arandaspis. To avoid multiplying nomenclature that no one will use anyway, let's refer to the psammosteidan variety as ps/aspidine.
 Like "normal" aspidine, ps/aspidine is three-layered. The basement layer is laminar, as in aspidine, and probably consists of dense, cortical bone, like some of the outer layers of endochondral bone. However, the basement layer is sometimes?) pierced by essentially vertical canals. Unlike aspidine, the middle layer is a thick spongy or trabecular space. It lacks the ordered columns of aspidine, although we are dealing with a difference in scale, and there may be less overall difference than meets the eye. A thin layer of cancellous bone left by a line of retreating or dying bone-forming cells is likely to look considerably more like trabecular bone if the layer is thick. In that circumstance, the osteocytes can overlap, slide past one another, and go off at angles. Furthermore, a thick layer is more subject to distortion from shearing and crushing forces, both during development and perhaps even after death. [3] Thus the middle layer of ps/aspidine differs from aspidine, but the degree of difference is unclear. Finally, the outer layer of ps/aspidine consists of dentine tubercles. According to Halstead, these dentine caps vary in some particulars from heterostracan dentine, but the differences do not appear to be large. As Halstead notes, the form of the dentine caps changes greatly on a specific level within Heterostraci, so one should probably not make too much of divergences between, for example, psammosteidans and pteraspidids.
Like "normal" aspidine, ps/aspidine is three-layered. The basement layer is laminar, as in aspidine, and probably consists of dense, cortical bone, like some of the outer layers of endochondral bone. However, the basement layer is sometimes?) pierced by essentially vertical canals. Unlike aspidine, the middle layer is a thick spongy or trabecular space. It lacks the ordered columns of aspidine, although we are dealing with a difference in scale, and there may be less overall difference than meets the eye. A thin layer of cancellous bone left by a line of retreating or dying bone-forming cells is likely to look considerably more like trabecular bone if the layer is thick. In that circumstance, the osteocytes can overlap, slide past one another, and go off at angles. Furthermore, a thick layer is more subject to distortion from shearing and crushing forces, both during development and perhaps even after death. [3] Thus the middle layer of ps/aspidine differs from aspidine, but the degree of difference is unclear. Finally, the outer layer of ps/aspidine consists of dentine tubercles. According to Halstead, these dentine caps vary in some particulars from heterostracan dentine, but the differences do not appear to be large. As Halstead notes, the form of the dentine caps changes greatly on a specific level within Heterostraci, so one should probably not make too much of divergences between, for example, psammosteidans and pteraspidids.
The interesting bit comes with the irregularities. To make a long story shorter, Halstead's microscopic examination of isolated plates demonstrated that damaged plates show evidence of extensive dentine regeneration. The photographic evidence, at least to the inexpert eye, seems very strong. The regenerated tissue takes the form, not only of new dentine tubercles, but also of pleromic, invasive dentine which seems to have filled gaps in the tissue. Halstead's microscopic evidence for regeneration, and thus for living cells, in the bone layers seems less convincing. However, the logic is certainly strong. If the dentine regenerated, it must have contained living cells capable of producing the tissue. These must have been connected to the circulatory system in some way absent some completely novel symbiotic arrangement). The most straightforward explanation is that, in spite of appearances, the circulatory system pierced the ps/aspidine; and, if so, there is no obvious reason why this peripheral circulation could not have maintained what Halstead calls aspidinocytes within the trabecular layer.
This is a murky area. Only one heterostracan, and no psammosteidan, shows evidence of the subaponevrotic vascular network that maintains a living integument in all other vertebrates. [4]. As noted, Halstead's own evidence for circulation, and for living cells within the bone framework, seems weak, particularly in comparison to the evidence for dentine regeneration. But, absent something really odd, is some sort of epidermal tissue peripheral to the bone. This seems even less satisfactory. Another odd bit of information noted by Halstead is that the specimens showing regeneration tend to be from specific sites. Conceivably, this geographic bias actually reflects some seasonal or other periodic process allowing a sort of temporary epidermis (with total resorption of the overgrown tissue). Or, we might be seeing a disease process common in some locations. Or, the evidence may reflect proliferation of an internal tissue triggered by traumatic breach of the exoskeleton under locally common conditions. Graduate students take note: these and other possibilities can probably now be examined. In fact, Halstead himself was a pioneer in, for example, the use of electron microscopy. However, the instruments and techniques now available are well beyond the reach of the technology reasonably available to Halstead in the 1960's. Paleohistology deserves a second look in the new millennium. ATW020126
[1] Psammosteids seem to have been restricted to the traditional "Old Red Continent:" the northern parts of Europe and Russia, various Arctic islands, Greenland, and much of North America. Tarlo (1964). Accordingly, there are no psammosteidans in the famous Australian Gogo Formation or the Devonian sites of China and Vietnam.
[2] Then again, a strong case could be made that Halstead's conclusion depends, in turn, on exactly what one means by acellular.
[3] As always, please recall that this is personal speculation and not to be taken too seriously.
[4] The exception is the cyathaspidiform Torpedaspis. Janvier (1986: 92, 94).
 Psammosteida: The largest and last of the pteraspidomorphs [JB93].
Psammosteida: The largest and last of the pteraspidomorphs [JB93].
Early Devonian to Late Devonian of North America, Russia & Europe
Protaspidoidea: Protaspididae + *. Drepanaspis + (Guerichosteidae + ((Pycnosteidae + Psammolepis) + Psammosteidae)).
 Characters:Medium to large (1m) forms [JB93] paddle-shaped [T64] with large, flat, almost round heads; plates similar to Pteraspididae, but with $ 2 pairs of orbital plates [H73]; orbit pierces anterior orbital plate; individual large plates separated from each other by small, polygonal tessellate platelets during ontogeny (young have no tessellations and large plates are contiguous); platelets capable of growth and have no central tubercle; terminal or subterminal mouth; small paired bone cornual "fins;" cornual plates reduced; branchial plates enlarged, flattened; branchial plates may be downturned in some forms as "outriggers" (?); branchial opening at posterior end of branchial plates [T64]; web-shaped caudal fins; $ trabecular "spongy") aspidine (cancellous layer lost -- see figure at aspidine); $ ornamentation of stellate, shiny tubercles; dentine tubercles very diverse in shape, density and size [T64]; dentine tubercles may vary in shape on different parts of same plate [T64]; dentine tubercles showing evidence of resorption & regrowth, thus probably covered with skin [H73]; presence of pleromic (gap-filling) dentine covering or even invading the bone in abraded or injured areas [H73]; sensory line in pore-canal system (?) [T64]; pores usually between tubercles, but may pierce ornament [T64]; sensory line pattern poorly known and may be asymmetrical [T64]; marine, presumably bottom-dwellers; later species restricted to deltas or floodplains.
Characters:Medium to large (1m) forms [JB93] paddle-shaped [T64] with large, flat, almost round heads; plates similar to Pteraspididae, but with $ 2 pairs of orbital plates [H73]; orbit pierces anterior orbital plate; individual large plates separated from each other by small, polygonal tessellate platelets during ontogeny (young have no tessellations and large plates are contiguous); platelets capable of growth and have no central tubercle; terminal or subterminal mouth; small paired bone cornual "fins;" cornual plates reduced; branchial plates enlarged, flattened; branchial plates may be downturned in some forms as "outriggers" (?); branchial opening at posterior end of branchial plates [T64]; web-shaped caudal fins; $ trabecular "spongy") aspidine (cancellous layer lost -- see figure at aspidine); $ ornamentation of stellate, shiny tubercles; dentine tubercles very diverse in shape, density and size [T64]; dentine tubercles may vary in shape on different parts of same plate [T64]; dentine tubercles showing evidence of resorption & regrowth, thus probably covered with skin [H73]; presence of pleromic (gap-filling) dentine covering or even invading the bone in abraded or injured areas [H73]; sensory line in pore-canal system (?) [T64]; pores usually between tubercles, but may pierce ornament [T64]; sensory line pattern poorly known and may be asymmetrical [T64]; marine, presumably bottom-dwellers; later species restricted to deltas or floodplains.
Links: ??????? Japanese); Stratený svet c.5/1999 (Czech); Drepanaspis gemuendenensis; Biology 356 (not really certain this is Drepanaspis, but its worth a close look).
Halstead 1973) [H73], Janvier & Blieck 1993) [JB93], Tarlo (1964) [T64].
Image of Drepanaspis model © North Eastern Geologic, courtesy of Gregory S. Gartland.
Note: [1] The structure and properties of aspidine (also spelled aspidin) have been the subject of a great deal of debate. We will tentatively proceed on the assumption that the labels are probably meaningless, although the structural details may be very significant. See Aspidine. [2] Some very small modifications were made to the figure from [T64] to clarify some of the divisions between plates. [3] Halstead and Tarlo are the same person. Tarlo changed his name in 1969 and moved from England (at least for a few years) to the University of Ife in Nigeria. Must be an interesting story there ... . ATW020112.
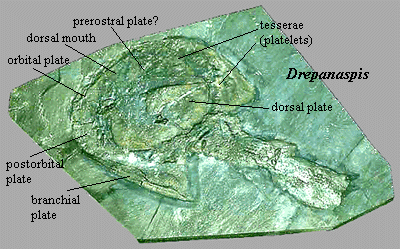 Drepanaspis: Drepanaspididae) An early, very well known, psammosteidan, "essentially a flattened Pteraspis." [H73: 294].
Drepanaspis: Drepanaspididae) An early, very well known, psammosteidan, "essentially a flattened Pteraspis." [H73: 294].
Range: lwD of Eur.
Phylogeny: Psammosteida: (Guerichosteidae + ((Pycnosteidae + Psammolepis) + Psammosteidae)) + *.
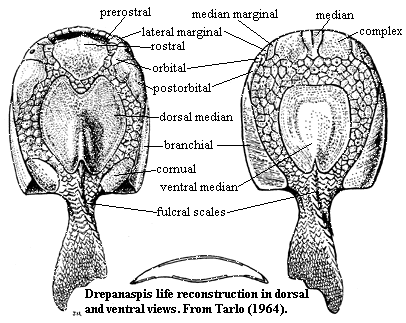
Characters: [3] dorsoventrally compressed and overall slightly convex body form [T64]; mouth dorsal [H73]; oral plates [2] large; 7 oral plates ornamented at terminal margins [H73], [T64]; oral plates fixed to anterior (ventral) margin of mouth [T64]; oral plates held inside mouth in life? [T64]; mouth with median tesserae just below ventral margin [H73] (contra, [T64]); thin, transverse prerostral plates across dorsal (= posterior) margin of mouth [H73]; small lateral marginal plate just anterior to orbital [T64]; small olfactory?) notch between lateral marginal & rostral [T64]; Y-shaped median plate on ventral mouth border, flanked by square complex plates and thin, sigmoidal median marginal plates [T64]; may be small pineal plate between rostral and dorsal median plates [T64]; dorsal median plate with medial indentation anteriorly, matching (but not meeting) posterior extension of rostral plate [T64]; orbital plate triangular [T64]; straight, vertical border between orbital and postorbital [T64]; long postorbital with irregular medial margin [T64]; postorbital articulates posterolaterally with branchial [T64]; ventral margins of lateral marginal, orbital and postorbital visible in ventral view [T64]; branchial plates long and narrow (primitive) [H73]; ventral shield mainly?) tesserae [H73]; branchial opening dorsal [H73]; in ventral view, posterior end of branchial plate transverse to body axis [T64]; broad tesserated zone between ventral mouth plates and ventral median plate [T64]; ventral median plate oval, but truncated anteriorly & posteriorly [T64]; median ventral with variably deep notch posteriorly [T64]; fulcral scales both dorsal & ventral [T64]; on both surfaces, posterior tesserae grade into caudal scales [T64]; tail almost equilobate [T64]; major plates show continuing concentric growth [H73] [T64]; ornamentation of small, stellate tubercles [T64]; marine [H73].
Links: Drepanaspis gemuendenensis; Biology 356.
References: Halstead 1973) [H73]; Janvier (1996) [J96], Tarlo (1964) [T64].
Note: [1] Earliest, but best known, psammosteidan. [2] "Oral plates" refers to the numerous, finger-like plates which were attached to the ventral surface of the mouth, either acting to cover it (perhaps as a coarse filter), or projecting anteriorly (perhaps as a scoop), or both. [J96]. As Halstead [H73] comments, the very fact that the oral plates are often found in articulation suggests that they were covered with flesh in life. Halstead [T64] also believes that the plates were held inside the mouth. [3] See figure at Psammosteida for plate nomenclature. 020111.
checked ATW060104