Pteraspidomorphi: Heterostraci 2)
Corvaspids and Cyathaspids
Taxa on This Page
- Corvaspididae X
- Corvaspis X
- Cyathaspidida X
Corvaspids: Mental Impressions and Ornamental Trees
Between the first Silurian appearance of Heterostraci and the Devonian radiation of definitive Cyathaspidiformes and Pteraspidiformes there is an impressively large group of poorly understood heterostracans: Athenaegis, cardipeltids, Tolypelepis, Traquairaspidiforms, and the Corvaspididae, to name some of the prominent elements of this jawless lumpenproletariat. As Janvier (1996: 95, 98) admits, a sensible phylogeny of the Heterostraci can be constructed only by ignoring these groups. This, however, strongly suggests that our current phylogenetic dispositions are perhaps not so sensible. At the moment (030511) we have reviewed only the corvaspids. Thus, we have little basis even for speculation. Nevertheless, the corvaspids do illustrate what some of the problems are.
The Corvaspididae have been around, under one label or another, since the 1930's. Much of the debate on nomenclature is, at bottom, no more than a pointless debate about taxonomic rank. For reasons discussed elsewhere, we decline to get involved in this issue. But, at the same time, it is difficult to arrive at a useful cladistic definition of the relevant group. We have, rather arbitrarily, put the corvaspids in with the Cyathaspidiformes. We have very little confidence in this assignment. The corvaspids might, as easily, be very basal pteraspidiforms or primitive Heterostracomorphi outside the Heterostraci. So, we can't define the corvaspids as a stem group because we don't really know what stem applies. Consequently, we are forced to apply an ad hoc crown group definition: "the last common ancestor of Corvaspis and Corveolepis and all of its descendants."
discussed elsewhere, we decline to get involved in this issue. But, at the same time, it is difficult to arrive at a useful cladistic definition of the relevant group. We have, rather arbitrarily, put the corvaspids in with the Cyathaspidiformes. We have very little confidence in this assignment. The corvaspids might, as easily, be very basal pteraspidiforms or primitive Heterostracomorphi outside the Heterostraci. So, we can't define the corvaspids as a stem group because we don't really know what stem applies. Consequently, we are forced to apply an ad hoc crown group definition: "the last common ancestor of Corvaspis and Corveolepis and all of its descendants."
What do we know about this tiny group? As it turns out, not very much. The dorsal armor may be a single plate, or may be comprised of as many as nine smaller plates. There may or may not be orbital plates. The ornamentation around the eye includes a unique and distinctive series of concentric rings of dentine, but nothing is known of the mouth or gill openings. The pineal area is located only a little behind the orbits, which is perhaps unusual for the Heterostraci. The main shields are extremely delicate towards their margins and tend to grade into smaller units -- particularly on the posterior margin of the dorsal plate. Here, the dorsal shield grades into the body squamation without a definite boundary, at least in some specimens. Since the armor margins are vague, we know little about the overall form of these animals. The armored area is certainly long and, at least by comparison with many other heterostracans, slim. But we cannot guess whether the body was round, deep or flattened. The tail is unknown; and, curiously, no fringing scales have been found. These large, angular scales, normally at the dorsal and ventral midline of the tail, are found in every other heterostracan. Furthermore, since the heterostraci had very little endoskeleton, fringing scales would appear to be necessary to hold the tail rigid enough to swim.
Essentially nothing is known of the internal structure. One photograph of an internal mold of the dorsal shield shows a "faint impression of the brain." Blieck & Karatajute- Talimaa (2001: 644, fig. 4B). This impression (shown at right) gives tantalizing hints of paired structures within and anterior to the medulla oblongata (where there ought not to be any) and a distinct, fan-shaped posterior expansion which is abruptly cut off. There are no impressions suggesting gill arches or lamellae, such as are found in many other heterostracan head molds. However, it is entirely possible that all of these features are artifacts of preservation or transmission, as we are looking at a scan of a photocopy of a photograph of a mold of an impression.
Finally, we arrive -- with much reluctance -- at the part of the journey where most schemes of heterostracan classification have come to grief: the ornamentation of the shield. The good thing about ornamentation is that essentially all early vertebrate fossils have it in abundance. If they did not, we would be unlikely to recognize them as vertebrate fossils. The bad thing is that ornamentation has a long and tortured history of phylogenetic misuse and misleading similarity. To understand how this happened, it is necessary to delve, briefly, into the mindset of science early in the last century. The scientific celebrities of the day, folks like Rutherford, Maxwell, Bohr and Einstein, were theoretical physicists. In practical matters as well, the first generations of adequately trained engineers were applying physical theory to transform the world in very obvious and tangible ways. The paradigm of theoretical development as the key to scientific understanding was transcendent. Zoologists and paleontologists were reluctant to be left off this bandwagon and sought to come up with theoretical approaches of equal breadth and explanatory power. This trend reached its apogee in the works of the Scandinavian school of Stensiö, Jarvik and, most especially, Bjerring (who pushed segmentalist theory almost past the point of absurdity) [1].
Against this background, it was entirely reasonable for Stensiö and others to construct an entire theory of vertebrate evolution based on changes in squamation. Accordingly, the older literature is full of abstruse terms used to describe the supposed development of "agnathan" integument based on equally theoretical long-term trends in the development of scale types. The problem is that squamation can't really tell us much. The basic structure of scales didn't change much during the Paleozoic. Blieck and Karatajute- Talimaa (2001) describe the trunk scales of corvaspids as rhomboidal, bearing parallel ridges of dentine posteriorly and flat-topped tubercles anteriorly (some of which may also bear ridges of dentine), with a small, unornamented anterior margin, and an anteriorly-protruding anchor process on the inner face. As noted below, this description matches scales of unknown Cambrian origin as well as it does the scales of Devonian corvaspids. It is easy to see that these units could be fused in various ways to form the surface ornamentation of various jawless fishes. However, this has relatively little to do with "tesserae," "synchronomorial units" or other, larger-scale organizational features, because the layer below the surface is made of acellular bone (aspidin). The aspidin layer may have some contribution from individual scales. Presumably, this is the source of the "compact aspidine" sublayer frequently seen in corvaspids and other Paleozoic fishes. However, the cancellous aspidine layer below is formed by a retreating front of dermal osteocytes, not by superficial cells associated with the squamation. Yet a third layer, the deep layer of laminar bone, is formed by yet a third developmental process, the superficial ossification of the basement membrane.
The point here is not that ornament and integument have no phylogenetic value, but that our information on these matters has been so thoroughly warped by scale-oriented theories that it is virtually impossible to sort out fact from theoretical construct. The actual patterns observed are probably rather complex results of the three independent developmental processes noted above, as well as taphonomic factors. A lot can happen to a fish in four hundred million years. Until someone is willing to go through the thankless task of re-analyzing about a century's worth of fish scale observations, we will be left with the decorative, but probably useless, ornamental trees of the Scandinavian school. ATW030511.
[1] Books could be written on the sociology of this movement and the efforts of the post-war generation of European scientists to overcome the dead weight of this paradigm. Phillipe Janvier's Early Vertebrates 1996) can, for example, be understood as the logical culmination of the post-war reaction. One of the greatest, but rarely recognized, accomplishments of cladistics has also been its influence in forcing scientists to focus once again on empirical investigation. We have argued elsewhere that empiricism can also be taken too far. The point made there, and here again, is that pragmatism should govern. Both empiricism and theory are only tools. Neither is an end in itself.
 Corvaspididae: Corvaspida of [S64]) stem group cyathaspids? [S64] includes the cyathaspid forms Archegonaspis & Poraspis in a "Corvaspiformes."
Corvaspididae: Corvaspida of [S64]) stem group cyathaspids? [S64] includes the cyathaspid forms Archegonaspis & Poraspis in a "Corvaspiformes."
Range: Pridoli to Early Devonian (Lochkovian) North America (Arctic Canada), Spitzbergen & Siberia Severnaya Zemlya) [BJ99]
Phylogeny: Cyathaspidiformes: (Cyathaspidida + Amphiaspidida) + *: Corvaspis + Corveolepis
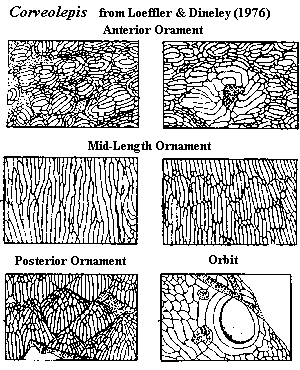
Characters: medium-sized [DL76]; dorsal & ventral plates each as single unit [BK01], but appears to be made up of fused scales [J96] (contra [S64]: may be two main dorsal plates) (and questioned [BK01]: dorsal plate either unitary or assemblage of up to nine plates, including paired orbital plates); orbits round and dorsolateral [BK01]; orbital plate ornamented with concentric ridges near the orbit, grading into short ridges & tubercles arranged in rings; orbital plate containing both supra- and sub-orbital elements (i.e., completely surrounding orbit) [S64] [DL76] (contra [BK01]: orbits completely surrounded, but perhaps by dorsal plate); branchial opening unknown [J96]; ventral plate tends to break up into smaller plates anteriorly [BK01]; ornamentation of wide, low, closely-spaced ridges with flattened surfaces and of tubercles with flattened peaks [S64] [DL76]; ridges short, smooth, grouped into units by network of grooves [LD76]; units may be separate or broken off main plates [BK01]; units, and margins of plates may be bounded by small, closely-packed tubercles [BK01]; posterior superficial shield units identical to isolated scales from area posterior to shield [LD76]; shield grades into posterior body squamation [LD76]; posterior body covered with small, rhomboidal scales [DL76] [BK01]; trunk scales with flat-topped crown, wide anterior shelf above thick, anteriorly-protruding anchor process [BK01] [3]; trunk scales with crown bearing wide longitudinal ridges with 1+ rows of anterior tubercles [BK01] [3]; ridge scales unknown and probably absent (unique among heterostraci) [BK01]; dentine ridges arise in clusters (??), each ridge being associated with 1-2 small pockets communicating with pulp canals [BK01] [4]; plate structure as shown in image, but some forms lack distinct compact aspidine layer [BK01] [4]; no actual tesserae as all fused together by basal layer of laminar bone [BK01]; sensory line system poorly developed or just poorly known, apparently a simple pit line system [BK01].
Note: [1] Stensiö uses the French word "ride" which translates literally as "wrinkle" or "ripple," both of which are more accurate descriptions than the usual English "ridge." On the other hand, who knows what he intended? Stensiö was a Norwegian who wrote in English, but is usually read in French translation. Today, he is best known through the critical work of Janvier, a Frenchman who usually writes in English. [2] Whether the orbits are formed by the dorsal plate or by a separate orbital plate, the fact that they are completely surrounded by one plate is considered diagnostic of the family. [3] Blieck & Karatajute-Talimaa [BK01] do not figure these scales. However, isolated scales are known which match this description rather closely -- from the Furongian! See [S98: 349]. [4] Note that the histology of the plates is quite similar to that of psammosteids.
Links: Upper Silurian and Devonian heterostracan pteraspidomorphs ( ...; 107.htm.
References: Blieck & Janvier (1999) [BJ99]; Blieck & Karatajute-Talimaa (2001) [BK01]; Dineley & Loeffler (1976) [DL76]; Janvier 1996) [J96]; Loeffler & Dineley (1976) [LD76]; Stanley 1998) [S98]; Stensiö 1964) [S64]. APW040316.
Corvaspis: C. kingi Woodward, 1934; C. karatajuteae Tarlo, 1965.
Range: Early Devonian (Lochkovian) of Europe & North America
Phylogeny: Corvaspididae: Corveolepis + *.
Characters: orbits as in Corveolepis? [LD76]; flat orbital plates [H73]; may have had separate median, orbital and ridge (or possibly branchial) plates [LD76] (contra? [H73]: single median plate with "occasional incorporation of tesserae at the margins"); arched branchial plates [H73]; usually interpreted as being broad and dorsoventrally compressed [LD76]; ornament with polygonal units with sculptured ridges [H73] [L95]; ridges of dentine [H73]; ridges of uniform height & width, with slight crenulation (i.e. a tendency to resemble grebeshkis) [S64]; tubercles of 2 kinds (a) rather thin & about the same width as the ridges, and (b) and thick, irregular ones located along the margins of the main plates [S64]; anterior portion of dorsal plate divided by grooves into several irregular polygonal fields, which are subdivided by other grooves into 2-5 fairly regular polygons ornamented with anteroposteriorly oriented ridges [S64] [2]; anterior and lateral margins with thicker tubercles [S64]; ventral disk also comprised of an epitega with tubercles marginally [S64].
Note: [1] From [H73: 293]: The "commonest ornamentation is of synchronomorial tesserae separated from each other by a narrow zone of tubercles. This suggests that the main tesserae were formed before the animal reached its definitive size..." [2]  See image at right from [S64]. Stensiö argues that these are cyclomorial scale units. The catch, to us, is that the strips of dentine line up perfectly from one "field" to the next, so that the same dentine ridge essentially continues from one unit to the next. Thus, it appears that the pattern of dentine ornament was superimposed on a pre-existing polygonal pattern of bone. The ridges could not possibly line up so perfectly if the dentine strips developed independently in separate cyclomorial scale units. In spite of Stensiö's insistence, the fields are not "assez régulièrement polygonaux." In fact, they are so irregular and so diverse that the dentine ridges simply could not line up in this manner from field to field if they were independently derived. Note also that the tubercles between fields also tend to form well-aligned strips over moderate distances at a 0-30° to the anteroposterior orientation of the superficial dentine ornament. Unfortunately for our own view, while Stensiö seems plainly wrong about the ornament in Corvaspis, his description is an almost completely accurate depiction of the ornament in Corveolepis. See [LD76] (and image below), [BK01]. This is odd, to say the least, since none of the specimens now known as Corveolepis were found much less described) until at least a year after Stensiö's review was published!
See image at right from [S64]. Stensiö argues that these are cyclomorial scale units. The catch, to us, is that the strips of dentine line up perfectly from one "field" to the next, so that the same dentine ridge essentially continues from one unit to the next. Thus, it appears that the pattern of dentine ornament was superimposed on a pre-existing polygonal pattern of bone. The ridges could not possibly line up so perfectly if the dentine strips developed independently in separate cyclomorial scale units. In spite of Stensiö's insistence, the fields are not "assez régulièrement polygonaux." In fact, they are so irregular and so diverse that the dentine ridges simply could not line up in this manner from field to field if they were independently derived. Note also that the tubercles between fields also tend to form well-aligned strips over moderate distances at a 0-30° to the anteroposterior orientation of the superficial dentine ornament. Unfortunately for our own view, while Stensiö seems plainly wrong about the ornament in Corvaspis, his description is an almost completely accurate depiction of the ornament in Corveolepis. See [LD76] (and image below), [BK01]. This is odd, to say the least, since none of the specimens now known as Corveolepis were found much less described) until at least a year after Stensiö's review was published!
Links: 107.htm; Untitled Document.
References: Halstead 1973) [H73]; Loeffler & Dineley (1976) [LD76]; Long (1995) [L95]; Stensiö (1964) [S64]. APW040316.
 Corveolepis: Blieck & Karatajute-Talimaa 2001. "Corvaspis" arctica Loeffler & Dineley, 1976; C. elgae Blieck & Karatajute-Talimaa 2001. Also includes, dubitante, Corvaspis cf. kingi, Corvaspis graticulata Dineley, and unnamed bits & pieces.
Corveolepis: Blieck & Karatajute-Talimaa 2001. "Corvaspis" arctica Loeffler & Dineley, 1976; C. elgae Blieck & Karatajute-Talimaa 2001. Also includes, dubitante, Corvaspis cf. kingi, Corvaspis graticulata Dineley, and unnamed bits & pieces.
Range: Early Devonian (Lochkovian) of Siberia Severnaya Zemlya) [B+02], Pridoli and/or Early Devonian of North America (Arctic Canada) [LD76], Spitsbergen.
Phylogeny: Corvaspididae: Corvaspis + *.
Characters: dorsal shield fused into single plate [BK01]; dorsal shield long, flat, & narrow [LD76]; smoothly rounded rostral margin [LD76]; dorsal shield completely encloses orbits [LD76]; orbits small, circular and anteriorly placed [LD76]; orbits directed dorsally, but located close to margin of dorsal shield [LD76]; orbits surrounded by annular ridges, and annuli by short, peripheral ridges [LD76]; annular orbital ornamentation grades into body ornamentation without sharp boundary [LD76]; pineal macula located slightly posterior to orbits [LD76]; dorsal plate lateral margins downturned to form ventrolateral laminae [LD76]; branchial openings not found or absent [LD76]; ventral shield unknown [LD76]; ornamentation of closely-spaced, smooth-topped dentine ridges [LD76]; ridges of uniform height & typically < 3mm long [LD76] [BK01]; ridge ornaments grouped into small, superficial units separated by grooves or rows of tubercles [LD76] [BK01]; subdivisions not present on rostral and lateral margins [LD76] [2]; anteriorly, units of ridges grouped around a single median ridge which is randomly oriented [LD76]; anterior ridges wider than elsewhere [LD76]; at midlength, ridges arranged variably into units, and orientation is uniform [LD76] [BK01]; posterior superficial units scale-like, grading into loosely attached units [LD76] [BK01]; loose rhomboid scales of posterior type associated with body fossils [LD76]; loose scales have unornamented anterior brim of 0.2 mm [LD76].
Note: [1] [LD76] described this form as a species of Corvaspis. The possibility exists that this may have affected the generality of their diagnosis and description. [2] This detail, as well as the superficial nature of the subdivisions, cuts slightly against any conclusion that all of the shield was assembled from true scales, as argued by, e.g., Stensiö [S64]. [3] [BK01] note that the actual shape of the body is unknown, as no specimens are free from possibly severe distortion.
Image: from [LD76].
Links: Untitled Document; Upper Silurian and Devonian heterostracan pteraspidomorphs ( ....
References: Blieck & Karatajute-Talimaa (2001) [BK01]; Blieck et al. 2002) [B+02]; Loeffler & Dineley (1976) [LD76]; Stensiö 1964) [S64]. APW040316.
 Cyathaspidida: streamlined forms with fusiform or cigar-shaped head-shields.
Cyathaspidida: streamlined forms with fusiform or cigar-shaped head-shields.
Entire body covered with plates and large, thick scales; anterior portion with single large dorsal & ventral shields; large plates may be formed from fusion of smaller units, fusing at maximal size; lateral branchial plate; seven pairs of gills; no known endoskeleton; large deep flange scales; caudal fin nearly symmetrical, but main axis may bend ventrally; no dorsal, anal, or paired fins; marine, later FW.
Image: Liliaspis from [L95]. Note the highly-developed sensory line system. Long identifies Liliaspis as an amphiaspid. He may be mistaken. The branchial plate seems to be separate, there is no tube-like rostral process as indicated in the figure legend, and the rostrum is broad and suggestive of a primitive pteraspidiform. Liliaspis found in the Urals, near the eastern extremity of the Old Red Continent, rather than from Siberia (from whence come all amphiaspids). It is more usually classified as a poraspid cyathaspidiform.
Links: Biology 356; Cyathaspid; Anglaspis; UALVP 17878 Cyathaspididae model; 106.htm; Museum Type Specimen.
References: Long (1995) [L95]. APW040316