Aves: Ratites: Struthioniformes
Taxa on This Page
- Aepyornithiformes X
- Apterygiformes
- Casuariformes
- Dinornithiformes X
- Dromornithiformes X
- Struthioniformes
The Icebird Suite: Part II
Flightless Phylogeny
| Neornithes
├─Tinamou
└─┬─Rhea
└─┬─Moa
└─┬─Ostrich
└─┬─Kiwi
└─┬─Cassowary
└─Emu |
Neornithes
├─Tinamou
└─┬─Moa
└─┬─Rhea
└─┬─Ostrich
└─┬─Kiwi
└─┬─Cassowary
└─Emu |
Neornithes
├─Tinamou
└─┬─Ostrich
└─┬─Rhea
└─┬─Kiwi
└─┬─Cassowary
└─Emu |
Rhea
Tinamou |
South America |
Cooper et al. (2001) |
Haddrath & Baker (2001) |
Harrison et al. (2004) |
Lithornis |
Europe,
North America |
mtDNA |
mtDNA |
mtDNA |
Ostrich |
Africa,
Europe? |
| Neornithes
├─Tinamou
└─┬─Lithornis
└─┬─Rhea
└─┬─Moa
└─Kiwi |
Neornithes
├─Tinamou
└─┬─Rhea
└─┬─┬─Cassowary
│ └─Moa
└─┬─Kiwi
└─Ostrich |
Neornithes
├─Tinamou
└─┬─┬─Moa
│ └─Kiwi
└─┬─Elephant Bird
└─┬─┬─Rhea
│ └─Ostrich
└─┬─Cassowary
└─Emu |
Elephant Bird |
Madagascar |
Bertelli & Chiappe (2005) |
Bertelli & Porzecanski (2004) |
Cracraft (2001) |
Cassowary
Emu |
Australia |
morphology |
morphology, mtDNA & RAG-1 |
morphology? |
Kiwi
Moa |
New Zealand |
Above are various recent family trees for the ratites, together with their known Recent distribution. We've also added Lithornis from the Eocene of Europe and North America.
All studies agree that the tinamou is basal, and we take that as established.
| Neornithes
├─Tinamou
└─┬─Ostrich
└─┬─Rhea
└─┬─┬─Elephant Bird
│ └─┬─Moa
│ └─Kiwi
└─┬─Cassowary
└─Emu |
Attempted consensus palaeognath phylogeny |
There is obviously very good agreement among the mitochondrial analyses, which is unsurprising, since they all use essentially the same data set. The frequently-cited work of vanTuinen et al. (1998) is not used because it looked only at a small number of taxa. Discarding that study, the first two mtDNA papers differ in only the branch order of rhea and moa (moa mtDNA isolated from fossils). The third study, Harrison et al. 2004), doesn't include moa DNA, and finds ostriches basal to rhea. We have very little faith in mtDNA studies as a general rule, for the reasons detailed here. However, the Harrison study is remarkable for its very high "bootstrap" values (a measure of internal consistency) and some special coding and data procedures which look, to us, to be a real improvement [1].
On the morphology side, we tentatively reject the combination approach of Bertelli & Porzecanski (2004). The idea of combining molecular and morphological data is, theoretically, a good idea. In practice, we don't think that anyone has yet worked out a good way to do it. Of course, our opinions are not to be taken very seriously. However, Robert Asher has probably worked harder on this area than anyone else, and his methods still (a) still require a number of very rough estimates to coordinate the data sets and (b) still come up with results which don't seem to be particularly likely. See Asher (2005). Bertelli & Porzecanski use a more straightforward method, but it seems not to address the problems which Asher has tried to solve.
 The remaining morphological studies both group moa and kiwi. That seems reasonable. A recent, unpublished morphological study (Dyke et al., 2004) agrees, and adds the elephant bird to this grouping, which is close to the result of Cracraft (2001). Having eliminated Bertelli & Porzecanski (2004), we also recover cassowary and emu as the terminal grouping in all studies which considered the two birds. The remaining two birds, rhea and ostrich, are usually found close together, but may be successive branches, rather than a separate group.
The remaining morphological studies both group moa and kiwi. That seems reasonable. A recent, unpublished morphological study (Dyke et al., 2004) agrees, and adds the elephant bird to this grouping, which is close to the result of Cracraft (2001). Having eliminated Bertelli & Porzecanski (2004), we also recover cassowary and emu as the terminal grouping in all studies which considered the two birds. The remaining two birds, rhea and ostrich, are usually found close together, but may be successive branches, rather than a separate group.
Putting this all together, the "consensus" cladogram is actually quite closely constrained, and would look something like the small cladogram in the chart above.
As discussed, (a) the moa/kiwi group might come before the ostrich/rhea step; b) ostrich [2] and rhea might be a clade, or might occur in the opposite order; and (c) the placement of elephant bird is not well supported. So this is not a completely determined cladogram. However, it's a whole lot better than the first time we tried this exercise, six years ago (in 1999).
At this point, the conscientious reviewer will roll out a series of molecular studies which purport to give the dates at which the ostriches split from rhea, etc. We decline to do so. Increasing evidence suggests that molecular clocks are simply not reliable. Don't just take our word for it: Heads 2005) provides numerous examples. And the combination of weird mtDNA phylogenies with eldritch mtDNA chronologies can yield results, published in serious journals, which are almost completely divorced from reality. See, e.g., Waddell et al. 1999). The best we can hope for there are some general ideas.
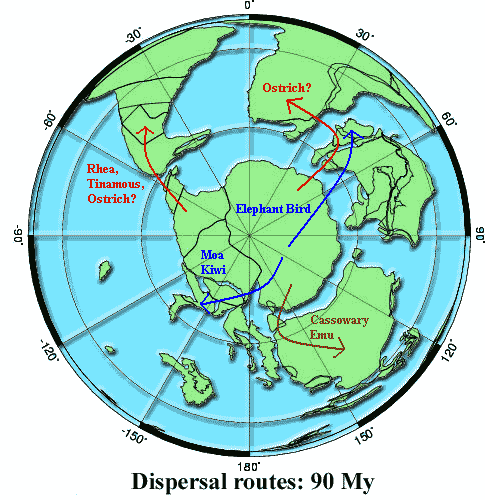
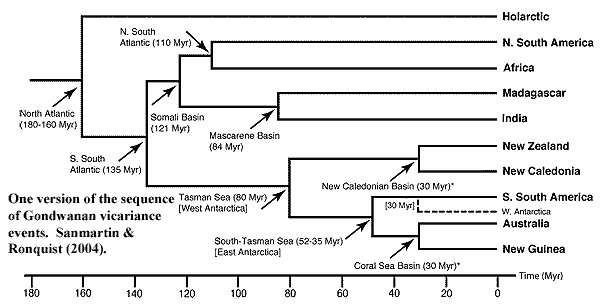
Getting Out of Gondwana
So where does all this leave us? Does vicariance work? The answer seems to be "almost." The figure shows the basic form of the dispersal from Gondwana over a South Polar map of about 90 Ma. How well does the trip report match the itinerary? From our previous discussion, you will hopefully have gained the sense that molecular phylogenies, much less molecular dating, are not yet a dependable source of information. And, even if we had correct dates, it's not clear when a land connection is really broken. Both sources of information should be carefully considered, but we can't depend completely on this sort of data. Finally, remember the California salamanders. Genetic "divergence" isn't always a simple concept. Ratites might have been a single, continuous breeding population from Argentina to Adelaide but still looked quite different at the two ends. Nevertheless, here is where the available information seems to be pointing.
Rhea and tinamous are non-problems. The path to South America was open for so long that timing is not really an issue. In fact, the connection to South America re-formed for some time after it was broken in the earlier Late Cretaceous. Tinamous are rather different from other paleognaths -- so much so that they are usually not considered to be ratites. Presumably, they evolved in South America quite early on. Rheas may date from the later land bridge, or at least from the same, presumably West Antarctic stock as the ostriches.
 The ostriches, once again, are the ones refusing to cooperate; and we don't yet have a really satisfactory explanation for them. Then again, perhaps the problem is too many explanations and not enough data. A northern route -- all the way through South and North America, across to Europe and south to Africa -- would explain their more northerly distribution and the embarrassing Paleotis, a bird from the Eocene (55 Ma) of Europe which looks suspiciously like an ostrich. Cracraft 2001). However, that route requires the ostrich stock to make a number of unlikely crossings, especially the big stretch (at that time) between South America and North America. In addition, we would have to suppose that ostriches on three continents became extinct (without leaving any known fossils except Paleotis), yet the African group somehow prospered. All this is possible, but the evidence just isn't there at the moment.
The ostriches, once again, are the ones refusing to cooperate; and we don't yet have a really satisfactory explanation for them. Then again, perhaps the problem is too many explanations and not enough data. A northern route -- all the way through South and North America, across to Europe and south to Africa -- would explain their more northerly distribution and the embarrassing Paleotis, a bird from the Eocene (55 Ma) of Europe which looks suspiciously like an ostrich. Cracraft 2001). However, that route requires the ostrich stock to make a number of unlikely crossings, especially the big stretch (at that time) between South America and North America. In addition, we would have to suppose that ostriches on three continents became extinct (without leaving any known fossils except Paleotis), yet the African group somehow prospered. All this is possible, but the evidence just isn't there at the moment.
Paleontologists have suggested a number of other routes for the ostriches. Sereno et al. (2004) have recently revived the idea of a transatlantic land bridge linking South America and Africa until as late as 95 Mya to explain some dinosaur vicariance problems. This is not too much of a stretch from more widely accepted dates beginning at 100 Ma. Cracraft (2001). Other possibilities with significant support include island hopping from Antarctica through India via the Seychelles (Rage, 2003), or via a temporary land bridge from India called the Kerguelen Plateau, or from temporary, volcanic islands created by the unsettled conditions around the India-Madagascar-South Africa confluence. Cracraft 2001). All of these may sound a little far-fetched. However, it only takes one pregnant female or a single breeding pair to establish a species, and a lot of very unlikely events can become almost inevitable on a timescale of 10-20 million years [3].
Alternatively, as Heads (2005) reminds us, we don't really have to assume that ostriches "arrived" in Africa at any particular time. They may well have been native to Africa -- just one link in a continuous chain of ratite species that circled Gondwana until it broke up. That may fit the facts best of all, since it wouldn't be completely out of line for a small breed of island-hopping ostrich to wander into Europe in the Eocene from Africa. However, the main exchange of species between Africa and Europe didn't begin until the Late Oligocene ~27 Ma) (Beresford et al., 2005), so we ought to be suspicious. What's worse, this hypothesis requires the whole ratite enterprise to start very early -- on the order of 120 Ma -- in order for ostriches to evolve in Africa while still maintaining some genetic connection with Antarctica. Again, we just don't have enough evidence to say at the moment.
The Moa-Kiwi-Elephant Bird complex is the most interesting part of the story. The other branches are located only on one continent, or on adjacent land masses. So, there's no telling if Rheas evolved in South America and never left, or whether they and ostriches both departed from West Antarctica. But Elephant Birds and Moas, in spite of a some strong physical similarities, literally lived on opposite ends of the Earth. Therefore, that group is really our top candidate for vicariance-driven evolution. That is, they are the ones most likely to have dominated the inland hills and cool forests of Old East Antarctica, the heartland of Gondwana, with the populations gradually moving apart as the South Pole became too cold to support large, flightless birds.
 Finally, the Cassowaries and Emus are closely related. They may have evolved on Australia, or arrived there from East Gondwana. Their overall similarity to ostriches and rhea may suggest the opposite ends of a ring species. However, the really close resemblance is between ostrich and emu. That may be more simply explained by the fact that both are extremely specialized for running on relatively flat ground. There are only so many ways to adapt to this lifestyle starting from the basic ratite body plan. So, it is not very surprising that they look much alike. For example, both have -- relative to their sizes -- perhaps the largest hindlimb extensor muscles of any tetrapod. Hutchinson 2004). To make best use of this running ability, both have evolved excellent vision based on heads placed well above ground level, relatively light, compact bodies, long legs, simplified feet, and other adaptations easily explained on functional grounds.
Finally, the Cassowaries and Emus are closely related. They may have evolved on Australia, or arrived there from East Gondwana. Their overall similarity to ostriches and rhea may suggest the opposite ends of a ring species. However, the really close resemblance is between ostrich and emu. That may be more simply explained by the fact that both are extremely specialized for running on relatively flat ground. There are only so many ways to adapt to this lifestyle starting from the basic ratite body plan. So, it is not very surprising that they look much alike. For example, both have -- relative to their sizes -- perhaps the largest hindlimb extensor muscles of any tetrapod. Hutchinson 2004). To make best use of this running ability, both have evolved excellent vision based on heads placed well above ground level, relatively light, compact bodies, long legs, simplified feet, and other adaptations easily explained on functional grounds.
Conclusion
So, our bottom line best guess -- much further out on a limb than most bird workers would care to perch -- is that there were two basic Gondwanan populations. The first was a sort of ring taxon of coastal or plains birds, with some original level of genetic continuity. The timing is still difficult to pin down, but these birds gradually split up and differentiated as the lines of genetic communication became more attenuated. The others, the moa-kiwi-elephant bird line, were inland, hill and forest types, who moved into the vacuum and away from the deteriorating environment. But this is largely speculation; and we will leave it with this image of the Last Gondwanan, somewhere in the coastal hills of East Antarctica, perhaps early in the Oligocene, weighing his options...
ATW051030.
Notes
[1] The improvements include (1) broad taxon sampling -- not within ratites, but in Neornithes generally, which is what the study was aimed at. In any case, because the shape of the neornithine tree is rather fuzzy, it makes sense to have many neornithine outgroups. Most mtDNA studies use the megapodes of Australia to root the tree, which is inadequate given the long branch lengths involved. (2) Third position codons were coded with an "RY" scheme, which seems not only a step in the right direction a priori, but also turns out to give better resolution than ACGT coding and partially corrects for codon bias (c.f. Phillips et al., 2001). (3) The coding scheme includes an element stating whether the nucleotide is contained in a stem (potentially double stranded region) or in a loop, with separate Gamma correction factors for stem and loop regions. The real novelty, and what seems to make the real difference, is RY coding, which is the feature unique to this study. The surprisingly strong effect of RY coding appears to be due to the significant differences in DNA base composition between bird lineages. These vary from
41% (swallow) to nearly 60% (whooping crane) (Edwards et al., 2002) so it appears that lack of correction for codon bias may strongly affect phylogenetic results.
 [2] One reason we prefer to put ostrich in the basal position is that several mitochondrial workers have complained that its "molecular clock" runs "too fast." That is, ostriches seem to have accumulated mtDNA mutations faster than they ought to have for their phylogenetic position. An alternative explanation, which we prefer, is that the clock runs just fine, thank you (or at least as well as those clocks ever run), but the phylogenetic placement is wrong. If ostrich is placed as the most basal ratite branch, then the clock result is more easily explained. Thus Harrison et al., who find ostriches branching early, have no complaints about ostrich clocks. Possibly this is because their clocks are set with multiple neornithine outgroups, such as -- that's right -- cuckoo clocks ...
[2] One reason we prefer to put ostrich in the basal position is that several mitochondrial workers have complained that its "molecular clock" runs "too fast." That is, ostriches seem to have accumulated mtDNA mutations faster than they ought to have for their phylogenetic position. An alternative explanation, which we prefer, is that the clock runs just fine, thank you (or at least as well as those clocks ever run), but the phylogenetic placement is wrong. If ostrich is placed as the most basal ratite branch, then the clock result is more easily explained. Thus Harrison et al., who find ostriches branching early, have no complaints about ostrich clocks. Possibly this is because their clocks are set with multiple neornithine outgroups, such as -- that's right -- cuckoo clocks ...
[3] Suppose that that the chance of a land bridge forming in a volcanic area is, on any given day, one in a billion. Is that a possibility we can ignore? On the contrary, over ten million years, the chances are about 97.5% that this event will occur at least once. Almost all evolution is driven by very unlikely events made inevitable by even longer stretches of time.
Struthioniformes:
Palaeotis, Struthio (ostriches).
Range: Modern forms from Early Miocene, but some fossils (Paleotis) attributed to this taxon from the Eocene.
Phylogeny: Ratites : (Rheiformes + ((Aepyornithiformes + (Dinornithiformes + Apterygiformes)) + Casuariformes)) + *.
Characters: Herbivorous two-toed African flightless birds. Largest living bird; head small, with lightly-built skull; neck long & mobile; wings strongly evident; $ 2 toes with loss of trochlea for 3rd on tarsometatarsus; feathers lack barbs; sexually dimorphic; live in groups of 10-50; live on fruits, nuts, other plant products in flat, arid country; very fast runners; male guards and incubates eggs.
Links: Ostrich Farming Database and Reference Library Index. (mostly ag); duke.usask ostr; Animal Diversity Web: Order Struthioniformes; World Birds Taxonomic List: Genera and species with citations.; Ostrich Struthio camelus); Struthioniformes; Struthioniformes Links; Struthio camelus; Birds: DNA Sibley's list - (Non Passeriformes). ATW010916
 Rheiformes: Rhea, Pterocnemia
Rheiformes: Rhea, Pterocnemia
Range: from the Eocene, South America.
Phylogeny: Ratites :: ((Aepyornithiformes + (Dinornithiformes + Apterygiformes)) + Casuariformes) + *.
Characters: tracheobronchial syrinx, relatively large wings; feathers on head, neck & thigh
Links: Rhea family peculiar courtship and incubation behaviors); Animal Diversity Web: Rheidae: Information; Rheiformes Mikko's Phylogeny); Rhea (bird) - Wikipedia, the free encyclopedia;
Aepyornithiformes:
Aepyornis (elephant bird).
Range: Pleistocene to Holocene of Madagascar.
Phylogeny: Ratites :::: (Dinornithiformes + Apterygiformes) + *.
Characters: Huge flightless birds, up to 3m and 450kg but most species much smaller). Graviportal, with thick, more short metatarsals; grazers or browsers; largest known bird egg at 30cm.
Links: Elephant Bird Egg from Madagascar; Roc historical and mythic); Greatest Places: Features: Big Egg: Links; The Egg Pages; Vorompatra Central (even better, see all of the links that Chip lists at Vorompatra Central: Other Sites).
 Dinornithiformes:
Dinornis (moas).
Dinornithiformes:
Dinornis (moas).
Range: Late Miocene to Holocene of New Zealand.
Phylogeny: Ratites ::::: Casuariformes + *.
Characters: Turkey-sized to very large (up to 3.3 m) extinct New Zealand ratites on the ostrich model. Diverse group, mostly at smaller end of size range; posture probable horizontal; no wings or pygostyle, tendonal canal in distal tibia similar to flying birds; gastroliths known.
Links: Moa Central; RSNZ Search Results (search on "moa"); ISLANDS.
 Apterygiformes:
Apteryx, kiwis.
Apterygiformes:
Apteryx, kiwis.
Range: from the Pleistocene of New Zealand.
Phylogeny: Ratites ::::: Dinornithiformes + *.
Characters: Chicken-sized flightless birds with long beaks. Long beak with nostrils near tip; bill has sensory bristles; eyes small and relatively weak in full daylight; uncinate processes present; fully ossified gastralia; embryonic kiwis have long reptilian tail; wings reduced to 5-10 cm rudiments (mentioned by Darwin); wing structure essentially normal; legs stout; feet with strong claws on all 4 toes; primary food items are worms & burrowing insects; functionally alternating ovaries; largest egg relative to size (~18cm long & 450g) with very high yolk (fat) content; nocturnal; male incubates as in other ratites.
Links: kiwi; Apterygiformes Links.
 Casuariformes:
Dromaius (emu), Casuarius (cassowaries).
Casuariformes:
Dromaius (emu), Casuarius (cassowaries).
Range: from the Late Oligocene of Australia.
Phylogeny: Ratites ::: (Aepyornithiformes + (Dinornithiformes + Apterygiformes)) + *.
Characters: Medium to large graviportal birds, 1-2 m, some with distinctive casque, from Australia and New Guinea. Boney forehead helmet ("casque") in forest-living forms; strong legs with claws; brown or black feathers with blue exposed skin, except wattles in bright colors; tail feathers absent; barbules absent on feathers; strong swimmers; leaping attack with claws; feed on fruits, berries, insects, small animals.
Image: of Dromaius © 1997 and reproduced with permission of Anderson's Creek Primary School.
Links: BIRDNET: Casuariformes. ATW000306.
Dromornithiformes:
Dromornis, Ilbandornis (mihirung).
Large to gigantic, somewhat like emus. Upper beak mobile on quadrate; probably not paleognathous; flat, ratite-like sternum; pelvic fenestra absent; typically reduced medial digit with hoof-like unguals; possibly forest herbivores; graviportal & cursorial forms. Sometimes classified as Casuariform, but may actually be basal galloanseriform.
Links: CNN Story; games.htm.