Tracheophyta
Vascular Plants: Maple > Moss
Introduction
 What we here call Tracheophyta is more exactly referred to as Pan-Tracheophyta. Cantino et al., (2007). As we will discuss, the correspondence is not exact. The definition we apply here is Quercus > Polytrichum, or maples > mosses. Generally speaking, the Tracheophyta are the vascular land plants and their morphologically similar, but avascular, cousins. They thus have a key position in the history of life.
What we here call Tracheophyta is more exactly referred to as Pan-Tracheophyta. Cantino et al., (2007). As we will discuss, the correspondence is not exact. The definition we apply here is Quercus > Polytrichum, or maples > mosses. Generally speaking, the Tracheophyta are the vascular land plants and their morphologically similar, but avascular, cousins. They thus have a key position in the history of life.
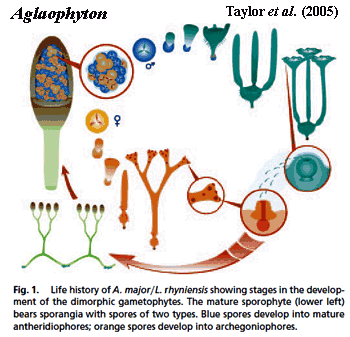
We can think of the Tracheophyta as including three sorts of plants: (a) a disorganized and grossly paraphyletic collection of basal "rhyniophytes," mostly from the Silurian and Early Devonian; (b) lycophytes and their relatives; and (c) the Euphyllophyta, a huge group made up of the ferns and woody plants. The coverage on this page deals with the rhyniophytes and only the most basal forms of the others. Elsewhere, we include individual pages on the rhyniophytes Horneophyton and Aglaophyton, as well as on two well-known, probably more derived, members of the group, Cooksonia and Rhynia.
The earliest fossil tracheophytes are of Wenlock age; but the clade probably diverged in the Llandovery -- possibly even the Late Ordovician. The history of photosynthesis on land keeps getting pushed back (see, e.g., Knauth & Kennedy, 2009), so its hard to be sure. Most of these basal forms disappeared by the end of the Devonian as seed plants (Spermatopsida) began to appear. Bateman et al. (1998).
Tracheophytes are often referred to as "vascular plants," which is close enough for most purposes. With few exceptions, tracheophytes have specialized water conducting elements (xylem) made up of dead cells (tracheids), with some kind of woody (or at least polyphenolic) reinforcement. Long strands of xylem conduct water because water is continuously removed from the upper parts of the plant by evaporation. This, in turn, occurs because water vapor escapes through pores, stomata. The presence of at least some stomata is thus likewise characteristic of tracheophytes. Sperry (2003). Actually, some non-tracheophyte groups have internal water-conducting elements, polyphenols, stomata, etc. But the phylogenetic distribution of these characteristics is spotty and does not point uniquely to one lineage as the sister group of tracheophytes. Ligrone et al. (2000).
An even better indicator of tracheophytes is their life cycle. Plants, you may recall, alternate between haploid (gametophyte) and diploid (sporophyte) generations. In all tracheophytes, the diploid, sporophyte phase of the life cycle is physically larger than the gametophyte, more or less independent of the haploid phase, and establishes direct contact with the substrate. Ligrone et al. (2000). This combination of features features seems to be truly unique to the Tracheophyta.
ATW090713. All text public domain. No rights reserved
Definition
Cantino et al., (2007) define Pan-Tracheophyta under Phylocode as "the total clade of Tracheophyta" and Tracheophyta as "The most inclusive crown clade containing Zea mays L. 1753 (Spermatophyta) but not Phaeoceros laevis (L.) Prosk. 1951 (Anthocerotophyta) or Marchantia polymorpha L. 1753 (Marchantiophyta) or Polytrichum commune Hedw. 1801 (Bryophyta)." Translated, this means everything more closely related to corn than to hornworts, liverworts, or mosses.
This definition is intended to promote stability. Cantino et al., (2007). Stability isn't usually that high on our agenda, and we prefer definitions that explicitly incorporate a specific hypothesis about the way things evolved. That usually works well enough for a platform like Palaeos. If we change our minds, we can go back and change everything. We have done just that, any number of times. However, our "maples > moss" definition (or "corn > moss" if you prefer) is more likely to be off base than most of the phylogenetic definitions we apply. For example, otherwise credible molecular phylogenies sometimes find that hornworts are more closely related to maples than to mosses. Nickrent et al. (2000); Qiu et al. (2006). If true, that would be awkward for us. And, certainly, few paleobotanists would clamber so far out on their respective limbs as to claim certain knowledge of the sister group to the tracheophytes. Nonetheless, we will stick to our short definition for the present.
ATW090713. All text public domain. No rights reserved
Phylogeny
 Tracheophyta marks an important and relatively sharp phylogenetic boundary. On the downstream side, the orderly and familiar succession of early tracheophyte clades are diplayed like the sunlit paths of a well-tended English garden. On the upstream side, we observe a dark and howling wilderness of mosses, liverworts, and hornworts in no comprehensible sequence or arrangement. We have -- somewhat arbitrarily -- chosen to cast our lot with the mosses as the sister group of Tracheophyta, at least for purposes of crafting a clade definition. That no longer seems as good a bet as it did just a few years ago. But no potential sister group seems capable of attracting a consensus of paleobotanists. Palmer et al. (2004). To be sure, paleobotanists are a flighty lot; but their lack of commitment seems amply justified in this case.
Tracheophyta marks an important and relatively sharp phylogenetic boundary. On the downstream side, the orderly and familiar succession of early tracheophyte clades are diplayed like the sunlit paths of a well-tended English garden. On the upstream side, we observe a dark and howling wilderness of mosses, liverworts, and hornworts in no comprehensible sequence or arrangement. We have -- somewhat arbitrarily -- chosen to cast our lot with the mosses as the sister group of Tracheophyta, at least for purposes of crafting a clade definition. That no longer seems as good a bet as it did just a few years ago. But no potential sister group seems capable of attracting a consensus of paleobotanists. Palmer et al. (2004). To be sure, paleobotanists are a flighty lot; but their lack of commitment seems amply justified in this case.
Kenrick & Crane's (1997) massive morphological study found that mosses were probably the closest living relatives of tracheophytes; but the conclusion was not strongly supported. The two main molecular studies have not added much clarity. Nickrent et al. (2000); Qiu et al. (2006). As mentioned above, they suggest that the hornworts may actually be closer, and that relationship is only slightly less parsimonious from a morphological perspective. Unfortunately, the DNA studies leaned heavily on chloroplast DNA. It turns out that, among the basal plants, rate heterogeneity is an even greater problem than in most organelle DNA. Soltis et al. (2002).
ATW090713. All text public domain. No rights reserved
The Consensus Phylogeny Within Tracheophyta
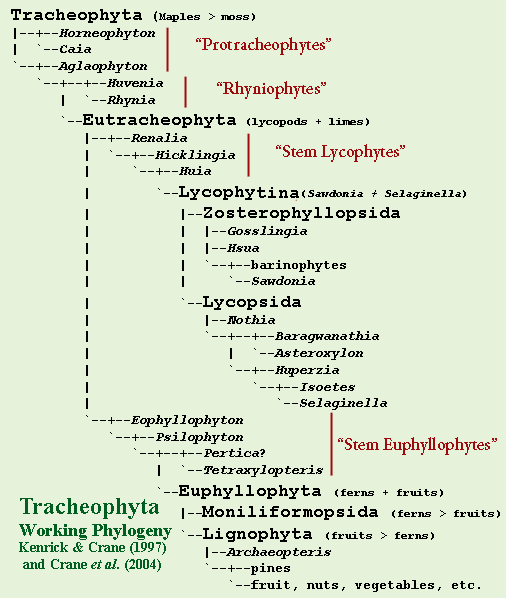
By contrast, within the tracheophytes, the main branches are remarkably uncontroversial. Magallón & Sanderson (2002) (chloroplast DNA); Kenrick & Crane (1997a) (morphology). With one exception, the outline determined by Kenrick & Crane (1997) has survived almost unchallenged.
According to this consensus view, the "protracheophytes" (e.g. Horneophyton, Caia, Aglaophyton), which lack xylem altogether, are the most basal tracheophytes. Kenrick & Crane (1997). Other well-known fossil plants from the Pragian Rhynie Chert ("rhyniophytes": e.g. Huvenia, Rhynia) are identified as more derived grades of the same basal tracheophyte stock. Of these, Rhynia may form a small clade with Stockmansella and Taeniocrada. Bateman et al. (1998). Cooksonia, known almost worldwide, often from much older deposits (Boyce, 2008), is said to be probably paraphyletic. Kenrick & Crane (1997a); Bateman et al. (1998). That is, some Cooksonia specimens are rhyniophytes, while others are believed to belong to more derived groups.
The term "pteridophyte" is sometimes used to refer to the paraphyletic group of all seedless Tracheophyta. The crown group of tracheophytes (limes + lycopsids) is the Eutracheophyta. This clade contains all living, and most fossil, vascular plants. Bateman et al. (1998).
 The lycophyte branch from this crown group presents some difficulties. The greatest difficulty is the problem of names. The taxon names Lycophyta, Lycophytina, Lycopodiophyta, Lycophytopsida, Lycopsida, etc. are used to mean very different things by different authors. We have been unable to find any real consistency -- even on our own website, much less in the literature. For our purposes we have made the following arbitrary choices.
The lycophyte branch from this crown group presents some difficulties. The greatest difficulty is the problem of names. The taxon names Lycophyta, Lycophytina, Lycopodiophyta, Lycophytopsida, Lycopsida, etc. are used to mean very different things by different authors. We have been unable to find any real consistency -- even on our own website, much less in the literature. For our purposes we have made the following arbitrary choices.
Lycophyta will be about the same as the Kenrick & Crane's (1997) Lycophytina or the Phylocode Pan-Lycopodiophyta: lycopsids > limes. It may turn out to include the same genera as Lycopsida. Then again, it may not. Tentatively, we follow Kenrick & Crane by placing Huia, Renalia, and Hicklingia inside Lycophyta but basal to Lycophytina. Crane et al. (2004). We will informally call this small, paraphyletic band of misfits "stem lycophytes." Huia is not to be confused with Hsua (a zosterophyll), an error which baffled us for some time. Not to mention Huperzia (an extant lycopsid) and Huvenia (a rhyniophyte).
Lycophytina doesn't have a close Phylocode equivalent. We use it to mean Sawdonia + Selaginella, that is, zosterophylls plus conventional lycopsids. This is an idiosyncratic use of this name. Don't even think about using it that way in public. However, we have to call this group something, and Lycophytina suggests a diminutive of Lycophyta -- which is exactly what we intend.
Lycopsida will mean Selaginella > Sawdonia, i.e., plants closer to living lycophytes than to zosterophylls. Again, Phylocode has no equivalent. To be completely honest, we envision Lycopsida as an intense, but somewhat homely teen-age girl with glasses, living in Queens. Zosterophyllopsida is her exotic, lesbian older sister from Paramus, New Jersey who drives a yellow Porsche. You will probably not find this to be a helpful aide memoire, but it may be worth a try. Almost anything is better than raw systematics.
Traditionally, the zosterophylls were thought to be just a basal grade of Lycophytina. Bateman et al. (1998). However, the current view is that zosterophylls are generally monophyletic and the sister group to an equally monophyletic Lycopsida. A few classical zosterophyll species are still of uncertain affinities. Crane et al. (2004). The zosterophylls are structurally simple, leafless plants, but they "strongly resemble lycopsids in sporangium morphology and in certain anatomical details." Examples: Sawdonia, Zosterophyllum, Huperzia. Kenrick & Crane (1997).
 The exact placement of basal, non-lycopsid Eutracheophyta is not well understood. Crane et al. (2004). Below Eutracheophyta, after some uncertain phylogenetic interval, we arrive at the crown group Euphyllophyta (a/k/a Euphyllophyta) = ferns + fruits. The stem group eutracheophytes -- below Eutracheophyta but above Euphyllophyta -- probably include early (mostly Late Devonian) fossils such as Eophyllophyton, Psilophyton, and Pertica. Bateman et al. (1998); Kenrick & Crane (1997: 129, fig. 4.33) (except as to Pertica); Schneider (2006). Stem euphyllophytes are sometimes called the "trimerophytes." Kenrick & Crane (1997).
The exact placement of basal, non-lycopsid Eutracheophyta is not well understood. Crane et al. (2004). Below Eutracheophyta, after some uncertain phylogenetic interval, we arrive at the crown group Euphyllophyta (a/k/a Euphyllophyta) = ferns + fruits. The stem group eutracheophytes -- below Eutracheophyta but above Euphyllophyta -- probably include early (mostly Late Devonian) fossils such as Eophyllophyton, Psilophyton, and Pertica. Bateman et al. (1998); Kenrick & Crane (1997: 129, fig. 4.33) (except as to Pertica); Schneider (2006). Stem euphyllophytes are sometimes called the "trimerophytes." Kenrick & Crane (1997).
The monophyly of Euphyllophyta is strongly supported by, among other things, "a unique 30-kb inversion in the chloroplast genome." Kenrick & Crane (1997a); Qiu et al. (2006) (criticized by Palmer et al., 2004, who argue that rare genomic events are no more privileged than rare morphological events). Euphyllophyta is, in turn, supported by two clades we will treat as stem groups: Monilophyta (ferns > fruits) and Lignophyta (fruits > ferns).
Monilophyta probably includes Equisetum (horsetails) as well as the ferns. The monophyly of the two was often doubted, but now seems increasingly accepted. Schneider (2006). The most basal known lignophyte is the famous Late Devonian forest-maker, Archaeopteris. Schneider (2006).
ATW090713. All text public domain. No rights reserved
A Misplaced Root?
 The main phylogenetic outline above is essentially undisputed. However, Boyce (2008) has raised a powerful dissenting argument about the position of the root. That is, even if the various groups are connected in the right way, we may be connecting them to the other Embryophyta in the wrong place. "Dissent" may be too strong a term to describe Boyce's position. Boyce is very cautious about drawing strong phylogenetic conclusions. However, he does point out some serious problems with the consensus structure.
The main phylogenetic outline above is essentially undisputed. However, Boyce (2008) has raised a powerful dissenting argument about the position of the root. That is, even if the various groups are connected in the right way, we may be connecting them to the other Embryophyta in the wrong place. "Dissent" may be too strong a term to describe Boyce's position. Boyce is very cautious about drawing strong phylogenetic conclusions. However, he does point out some serious problems with the consensus structure.
First, as everyone has known for many years, there's a timing problem. The protracheophytes and rhyniophytes are all Early Devonian except for Cooksonia. Yet -- despite many attempts to discredit the evidence -- undoubted Lycophytina are known from the Wenlock. Kotyk et al. (2002). That leaves a very long ghost lineage of protracheophytes, rhyniophytes and assorted stem Lycophyta. This is not impossible, but it is unsettling. Second, Devonian species of Cooksonia appear to eutracheophytes. One species in particular appears to be a zosterophyll (although it lacks a Porsche). This is also possible, if also disturbing. Cooksonia may not actually be a monophyletic genus. It may consist of a group of unrelated plants, or it may be grossly paraphyletic. That is, all eutracheophytes may be actually descended from Cooksonia.
But here's the real kicker. Early Cooksonia (or "cooksonioids," including Cooksonia, Salopella, Tarrantia, Tortilicaulis) have stems which are 100µ or less in diameter. Boyce shows that this is simply too skinny to support both vascular and photosynthetic systems. The combination requires a minimum stem diameter of about 700µ -- much stouter than Silurian Cooksonia. It is hard to imagine a plant the height of Cooksonia (several cm) without a water transport system. Even the miniature sporophyte axes of mosses generally have dedicated water transport systems. Boyce (2008).
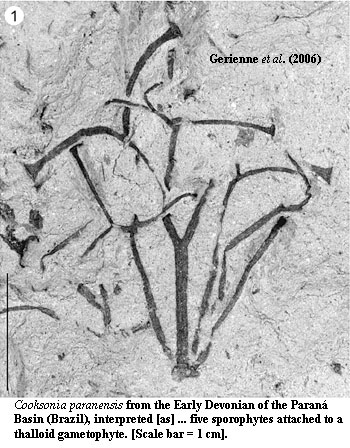
Therefore, the implication is that Cooksonia lacked enough photosynthetic capacity to be self-sustaining. If so, Silurian Cooksonia were not independent sporophytes, as is usually supposed, but were "parasitic" on a gametophyte -- just like the sporophytes of mosses, liverworts, and other early embryophytes. This supposition is confirmed by other evidence: the similarity of Cooksonia proportions to other non-photosynthetic stalks, the low density of stomata, the presence of a supporting cortical sterome (taking up even more of the limited space in the stem), and the absense of roots or root analogues. If correct, then Cooksonia is the otherwise missing link between early plants and true tracheophytes. That is, it is clearly a large (by Silurian standards) sporophyte, which looks like a tracheophyte, but was not fully independent. Boyce (2008). Recently Gerienne et al. (2006) have reported a very large specimen of Cooksonia which they tentatively interpret as five sporophytes emerging from a thalloid gametophyte.
 If Boyce is correct, we should redraw the phylogeny of tracheophytes with the same branches, but with the root moved to look more like this:
If Boyce is correct, we should redraw the phylogeny of tracheophytes with the same branches, but with the root moved to look more like this:
Embryophyta
├─Bryophyta
└─Cooksonia
├─┬─"Rhyniophtes"
│ └─"Protrachophytes"
└─┬─┬─Zosterophyllopsida
│ └─Lycopsida
└─Euphyllophyta
 Or, since some species of Cooksonia are very zosterophyll-like:
Or, since some species of Cooksonia are very zosterophyll-like:
Embrophyta
├─Bryophyta
└─Cooksonia
├─Zosterophyllopsida
└─┬─Lycopsida
└─┬─┬─"Rhyniophtes"
│ └─"Protracheophytes"
└─Euphyllophyta
Actually, we are quite fond of this last one. First, it comes closest to explaining why we find Cooksonia and lycopsids in the Silurian, rhyniophytes in the Early Devonian, and euphyllophytes only later. The thesis does require secondary simplification in rhyniophytes, but this is entirely possible. In fact, it has long been suspected that the Rhynie Chert plants are secondarily simplified as an adaptation to a very unusual, wet, acid environment. See, e.g., Bateman et al. (1998); Arnold (1947: 80) (referring to even earlier work). Psilotum looks very much like a rhyniopsid, but is actually a living, secondarily simplified euphyllophyte.
Second, Boyce's thesis also explains why Cooksonia species often distribute themselves so widely in most cladograms. The genus appears to be paraphyletic -- not because it is a garbage taxon -- but because it is paraphyletic. Ultimately, the only reason we have not adopted this revisionist phylogeny is that Boyce himself has chosen not to assert it directly.
ATW090713. All text public domain. No rights reserved.
Diversity and Occurrence
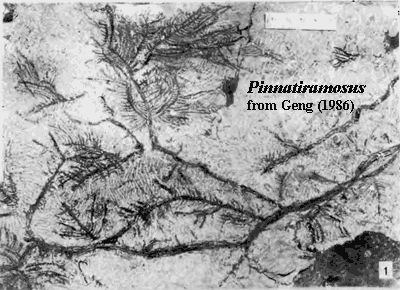
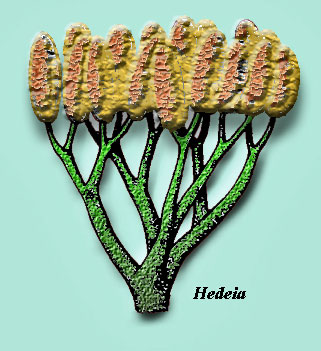 Tetrahedral spores (suggesting haploid cells produced by meiosis) with decay-resistant walls (presumably including sporopollenin) are diagnostic of land plants. These spores appear in the fossil record as early as the Middle Ordovocian. Kenrick & Crane (1997a); Bateman et al. (1998). Tracheophyte body fossils are not known before the Wenlock. However, Silurian tracheophyte fossils already include lycopsids (like Baragwanathia, pictured above) and zosterophylls, as well as the less-easily classified Salopella, Hedeia, and the amazingly advanced -- but equally controversial -- Pinnatiramosus (Geng, 1986).
Tetrahedral spores (suggesting haploid cells produced by meiosis) with decay-resistant walls (presumably including sporopollenin) are diagnostic of land plants. These spores appear in the fossil record as early as the Middle Ordovocian. Kenrick & Crane (1997a); Bateman et al. (1998). Tracheophyte body fossils are not known before the Wenlock. However, Silurian tracheophyte fossils already include lycopsids (like Baragwanathia, pictured above) and zosterophylls, as well as the less-easily classified Salopella, Hedeia, and the amazingly advanced -- but equally controversial -- Pinnatiramosus (Geng, 1986).
Pinnatiramosus is controversial because it has a number of features which seem to have no business on a plant from the Llandovery. For example, it has well-developed pitted tracheids and fairly regular, plainly monopodial branching. Geng (1986). The branching pattern is particularly startling because this kind patter of development requires lateral meristem. As we will see, that is unexpected in a plant from the Silurian. The ultimate order of branch is 400-800 µ in diameter -- just possibly stout enough to be a fully developed photosynthetic structure. Boyce (2008). Pinnatiramosus does not appear to bear leaves of any sort, nor even enations; and no reproductive structures are described. Geng (1986). Some relatively recent work (which we were not able to lay our hands on) argues that Pinnatiramosus is actually the intrusive root system of an unidentified plant from overlying deposits of Permian age.
On the other hand, there is quite solid evidence of lycophytines of various sorts from Ludlow exposures in both Australia and Canada. In addition to the lycopsid Baragwanathia, the Australian Ludlow flora includes Salopella, Hedeia, and zosterophylls. Kotyk et al. (2002) have documented an extensive zosterophyll fauna from the Ludlow of arctic Canada. This suggests that tracheophytes diverged from other embryophytes in the Llandovery or even the Late Ordovician. Thus, by the time that plant fossils become abundant in the Early Devonian, the pace of plant evolution -- while relatively brisk -- could not quite be described as explosive. Kenrick & Crane (1997a).
 The genera treated as the most basal tracheophytes in the consensus phylogeny (e.g., Aglaophyton and Horneophyton) are referred to as "protracheophytes" because they lack tracheids. Bateman et al. (1998). However they don't appear in the fossil record until the Early Devonian. This is the age of the remarkable Rhynie Chert fossils, which have structures perfectly preserved at the cellular level. "Unfortunately, the supposedly archetypal Rhynie Chert flora appears to be an unusual assemblage specialized for life in a low-pH, periodically flooded habitat; the flora may have included species secondarily reduced for aquatic life habits." Bateman et al. (1998).
The genera treated as the most basal tracheophytes in the consensus phylogeny (e.g., Aglaophyton and Horneophyton) are referred to as "protracheophytes" because they lack tracheids. Bateman et al. (1998). However they don't appear in the fossil record until the Early Devonian. This is the age of the remarkable Rhynie Chert fossils, which have structures perfectly preserved at the cellular level. "Unfortunately, the supposedly archetypal Rhynie Chert flora appears to be an unusual assemblage specialized for life in a low-pH, periodically flooded habitat; the flora may have included species secondarily reduced for aquatic life habits." Bateman et al. (1998).
Euphyllophytes (ferns and lignophytes) appear in the Late Devonian. By the end of the Devonian, well-developed forests of the basal lignophyte Archaeopteris covered broad areas of lowlands. Thus morphologically identifiable representatives of all the main tracheophyte groups all appear in the fossil record over a period of only about 70 million years, during the late Silurian and Devonian. Bateman & DiMichele (2003).
ATW090714. All text public domain. No rights reserved.
uploaded ATW050928
modified ATW090714

