
| Metazoa | ||
| Metazoa | Metazoa |
Eukarya
└─► |
|
Life
├─Eubacteria
└─┬─Archaea
└─Eukarya
├─Chlorobionta
└─┬─Fungi
└─Metazoa
├─Choanoflagellata
└─Porifera
├─Radiata
└─┬─Cnidaria
└─Bilateria
├─Deuterostomia
└─Protostomia
├─Ecdysozoa
└─Spiralia
|
Quick Links to Major Animal Groups |
Editor's note
This page is in the process of being revised, but is posted as is for now as a holding page and to provide links to other pages. Because Palaeos is still under construction and major revision, not all links given here work. MAK120115
Arthropods: insects, crustaceans, scorpions -- the most successful metazoans.
Brachiopoda: they look like mollusks, but they're not. 
Bryozoa: advanced, encrusting reef-builders. ![]()
Chordata: mostly, the Vertebrates. 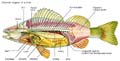
Cnidaria: Radiate animals -- jellyfish and corals. 
Echinoderms: our cousins, the sea urchins, sand dollars, starfishes and so on. ![]()
Mollusca: clams, oysters, and the like. ![]()
Porifera: sponges. 
Halkieriida sometimes bizarre worms near the root of the Spiralia. ![]()
Tardigrada: "water bears." 
 The Animal Kingdom (Metazoa) is usually considered to include multi-cellular, heterotrophic eukaryotes in which (unlike Plants) the cells are without cell walls. Everything from sponges and jellyfish to insects and vertebrates is belongs in "Metazoa", and considered to have evolved from a single unicellular choanoflagellate ancestor, sometime during the Ediacaran period.
The Animal Kingdom (Metazoa) is usually considered to include multi-cellular, heterotrophic eukaryotes in which (unlike Plants) the cells are without cell walls. Everything from sponges and jellyfish to insects and vertebrates is belongs in "Metazoa", and considered to have evolved from a single unicellular choanoflagellate ancestor, sometime during the Ediacaran period.
From a simple colonial choanoflagellate, animals developed through increasing grades of specialization and complexity: first sponges, then coelenterates, and finally bilateral animals possessing a head and distinct right and left sides). A recent interpretation of this monophyletic animal kingdom theory is the phylogenetic scheme of Wainright et al. 1993 shows choanoflagellates contained within the monophyletic assemblage Metazoa (= "animals"), and Fungi as the closest sister group to Metazoa. We thus conceive of Metazoa as the sister group of the Fungi, and use the term phylogenetically to mean toads > toadstools.
The problem here is that, although choanoflagellates seem clearly related to sponges, it is not clear how closely related sponges are to the rest of the Metazoa. It is also difficult to see how such a locally organized organism as a sponge (essentially nothing but a glorified colonial protozoan) can develop into an organism with a specific body structure and internal organs. The most widely held theory seems to be that a colonial choanoflagellate evolved into a hollow spherical ball of cells, the blastula, which constitutes the earliest embryonic stage of development, and even occurs in sponges. The 'blastula model' of metazoan evolution goes back to the famous 19th century German Darwinist Ernst Haekel.
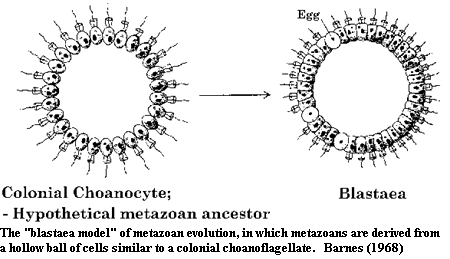 However, it is not certain that such a blastaea animal ever even existed. The theory that "ontogeny recapitulates phylogeny," championed by Haekel (according to which the growing embryo passes through all its past evolutionary stages - e.g. the early human fetus possesses gill slits and a tail), was enormously popular for some time.
However, it is not certain that such a blastaea animal ever even existed. The theory that "ontogeny recapitulates phylogeny," championed by Haekel (according to which the growing embryo passes through all its past evolutionary stages - e.g. the early human fetus possesses gill slits and a tail), was enormously popular for some time.
All we can say for certain is that, some time during the Late Proterozoic era, an unknown protozoan (or protistan) organism developed into a tiny colonial form, which eventually became the common ancestor of the Metazoa. The actual nature of this organism is not known, as it was soft-bodied and left no trace. It used to be thought that sponges evolved from a different single celled organism to higher animals (in which case the Metazoa are a polyphyletic taxon), but recent molecular phylogenetic evidence indicates this is not the case.
The base of the animal tree is moderately non-controversial. Animals are closely related to a particular group of protists, the choanoflagellates. The first metazoans were probably sponges, Porifera. Sponges are multicellular animals with specialized cell types, but no specialized tissues. The different cell types are intermingled, and one part of a sponge looks rather like another. The Porifera are probably paraphyletic. That is, all other animals are probably descended from something we would probably think of as a "sponge." In that sense, all animals may be members of the Porifera. However, we will use the term only as it applies to sponge-like animals. More generally, animals without strongly differentiated tissues are sometimes referred to as Parazoa, since there are probably animals other than sponges with this grade of organization, e.g., the Archaeocyatha and the Ediacaran fauna.
All animals with distinct tissue types are referred to as the Eumetazoa. Phylogenetically, we might define the Eumetazoa as the crown group uniting jellyfish and silverfish. The jellyfish lineage includes the Cnidaria jellyfish, corals, etc.) and some related types. These animals, the Radiata, seem to have derived from a single common ancestor who was not in our direct line of descent. The Radiata all have differentiated tissues, but normally nothing much more complicated than "inside" (endoderm) and "outside" (ectoderm), with some level of front-to-back specialization. From an embryological standpoint, they lack mesoderm, a tissue type which characterizes all more derived animals. Most are radially symmetrical. Their body plan is based on a cylinder open at one end, with the open end serving both for ingesting food and eliminating waste.
All other animals are Bilateria. Bilaterians undergo a more complex gastrulation and possess mesoderm. Thus, they begin with three embryonic cell types. Furthermore, they are bilaterally symmetrical. In addition to a more complex inside-outside pattern of tissues and the ancestral front-to-back organization, Bilateria have a separate top and bottom, with symmetrical left and right sides. In many worm-like forms, this top-and-bottom asymmetry is not well developed on the outside. However, the internal organization generally involves distinct dorso-ventral organization, with, for example, muscles and circulatory structures located dorsally, and the gut and a major nerve chord located ventrally. Most (but not all) also have a separate mouth and anus, so that the flow of digestion is one way.
At this point, things become much less clear. There are three groups of Bilateria which show some internal cohesion. These are the Deuterostomia including echinoderms and chordates), the Spiralia including annelid worms and mollusks), and the Ecdysozoa (including arthropods). Each of these groups includes some of the "minor phyla" of animals. However, a large number of mostly worm-like groups are left out of this scheme; and their positions are sometimes too vague even to guess at.
Of the three main groups, the deuterostomes probably branched off earliest, but even this has been disputed. The term "protostome" is used a good deal, particularly in the older literature, to refer to the lophotrochozoans, ecdysozoans, and everyone else who exhibits a particular pattern of early embryonic development. For our purposes, we will treat the deuterostomes as the earliest-branching clade and use the term "protostome" to refer to the crown group of bugs + slugs (Ecdysozoa + Lophotrochozoa). Other than the most well-established members of the two protostome groups, all other animal phyla will be treated as Bilateria incertae sedis.
With that said, our tree looks like this:
See the dendrogram page for a more detailed coverage.
Most of the "Halkeriids" are probably closer to mollusks than to annelids.
ATW041229. Slightly revised ATW070725. Text public domain. No rights reserved.
Metazoa Haeckel 1874 = Animalia
Range: Fr Ediacaran
Phylogeny: Eukarya ::: Stem Metazoa :Fungi + * : paraphyletic Porifera ::: Eumetazoa
develop from a blastula, cellular to organ-systems grade, food ingesting without chloroplasts, subdivided on grade of organization, symmetry, and coelomic development
Comments: The Animal Kingdom. Strictly speaking, Animalia Linnaeus 1758 has priority. However on the one hand "animal" tends to be associated in the minds of the average person with vertebrates, , and specifically, mammals , whereas "metazoan" is a nice, neutral-sounding technical term, free of colloquial nuances. In older science textbooks Kingdom Animalia traditionally also includes the heterotrophic (animal-like) protsits. Hence we have adopted Metazoa defined in terms of Whittaker & Margulis's five kingdom model, with multicelluarity as the arbitrary dividing line between paraphyletic protista and multicelluar animals. MAK120423
page uploaded 7 April 2002; last modified ATW070725, checked ATW070725; last modified MAK120421

