illustration © Prof. Silvio Renesto, University of Milan - Vertebrate Palaeontology
| Late Triassic epoch | ||
| Mesozoic: Triassic Period |
Norian Age - 2 |
| Carnian | Middle Triassic | Norian 1 | ||
| Rhaetian | Early Jurassic | Norian 3 | Timescale |
As for actinopterygian fish, the Late Triassic saw the first real radiation of Neopterygii, a radical ecological shift equivalent to the ascendancy of Teleosts during the Cretaceous. Significantly, more primitive forms - Chondrosteans and "Sub-Holosteans" - remain important, especially at the top of the food chain. In localities like the richly fossilferous Zorzino Limestone (Calcari di Zorzino), where at 50 genera have been described, all the large predators (Birgeria, Saurichthys) are chondrostean [ref Prof. Andrea Tintori, Norico Fauna and Flora ]. The following are some representative Actinopterygia of this time. All the species illustrated here were contemporary, and lived in the European Tethys, during the middle of the Norian.
|
|
illustration © Prof. Silvio Renesto, University of Milan - Vertebrate Palaeontology |
Subclass: Actinopterygii
Infraclass: Neopterygii
Division: "Holostei"
Order: Semionotiformes
Family: Dapediidae (or Semionotidae)
body length of 8-9 cm
Zorzino Limestone
Norian
Zogno, Italy (tropical Tethys)
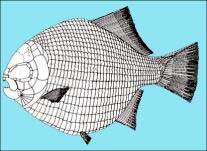
A common deep-bodied late Triassic and Early Jurassic durophagous fish. D. noricum is the smallest known species of the genus, it also differs from other species in the shape of the infraorbitals.
Dapedium by Prof. Andrea Tintori (also link to larger illustrations on this page)
 |
|
illustration © Prof. Silvio Renesto, University of Milan - Vertebrate Palaeontology |
Subclass: Actinopterygii
Infraclass: Neopterygii
Division: "Holostei"
Order: Semionotiformes
Family: Semionotidae
body length from 30 cm to 1 meter
Zorzino Limestone; Dolomia di Forni
Norian
Brembilla; Endenna; Poscante; Cene; Valvestino; all Northern Italy (University of Milan - Vertebrate Palaeontology collection - see Norian localities page), also known from elsewhere in Europe (teeth of similar or same species also known from the Norian-Rhaetian of Europe, especially from bone-beds of England, France and Germany) (tropical and subtropical Tethys)
The "big brother" of Dapedium, Sargodon was the largest of a number of species of durophagous (feeds on hard shelled food) or semi-durophagous fish of this region and time
Sargodon by Prof. Andrea Tintori (also link to larger illustrations on this page)
 |
|
illustration © Prof. Silvio Renesto, University of Milan - Vertebrate Palaeontology |
Subclass: Actinopterygii
Infraclass: Chondrostei
Order: Luganoiiformes
Family: Thoracopteridae
Carnian of Raibl (Italy) and Lunz (Austria), also known from the Norian of Italy (Tethys)
The earliest known flying (actually, gliding) fish, and the only type known before the Cenozoic exocetids, with which they shared a similar lifestyle.
Norico Fauna and Flora by Prof. Andrea Tintori
 |
|
illustration © Prof. Silvio Renesto, University of Milan - Vertebrate Palaeontology |
Subclass: Actinopterygii
Infraclass: Chondrostei
Order: Saurichthyiformes
Family: Saurichthyidae
the largest Saurichthys individuals from the Zorzino Limestone attain a body length of 180 cm
Norian of the Southern Calcareous Alps
An ubiquitous form, known worldwide from the Early to Late Triassic, Saurichthys was the Triassic equivalent of a pike or barracuda. By Norian times, perhaps in response to selection pressure by the nimble newly emerging Neopterygia, a number of advanced species of this genus had evolved, distinguished by a highly stiffened vertebral column, resulting in a greater springiness and hence faster acceleration. An ambush predator.
Saurichthys by Prof. Andrea Tintori
 |
|
illustration © Prof. Silvio Renesto, University of Milan - Vertebrate Palaeontology |
Subclass: Actinopterygii
Infraclass: Chondrostei
Order: Palaeonosciformes
Family: Birgeriidae
Early to Late Triassic
Spitzberg, Greenland, North America, Madagascar, Europe (Early Triassic)
Like Saurichthys, Birgeria was a large primitive form that nevertheless survived well as a dominant predator for a considerable period of time
 |
|
illustration © Ron Garrett, used with permission |
The warm Triassic seas are home to a rich marine fauna. Reefs of brilliantly coloured corals and sponges provide shelter for fish, Crustacea, and other animals. Rising upon the water column of the open seas we find the trophic food chain that has supported higher life on Earth for hundreds of millions of years. Warm sunlight nurtures phytoplankton, which supporting millions of tiny zooplanktonic organisms, which provide food for small fish.
These fish are in turn preyed upon by larger fish and the coiled-shell ceratite ammonoids and squid-like coleoid cephalopods. These in turn fall victim to the snapping jaws of small to medium-sized ichthyosaurs, marine reptiles not unlike modern dolphins, and averaging some 3 meters in length. But even these ichthyosaurs are not the top of the food chain. For ruling the Triassic waves in the earliest Norian, as it did in the late Carnian and will continue to do so throughout the Norian, is the mighty Shonisaurus, a whale-sized predator on anything smaller than itself. For a span of at least fifteen million years - from early Late Carnina to Late Norian and maybe beyond, these ichthyosaurs - far larger than any Cretaceous T. rex - will continue with only minor (species-level) change.
|
|
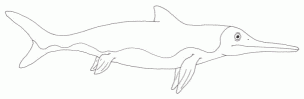 |
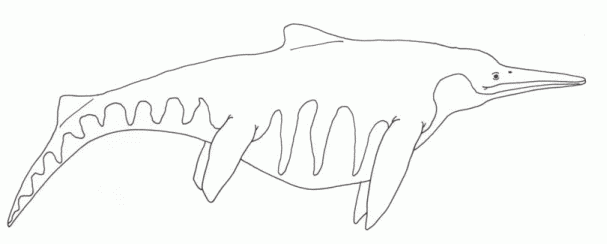
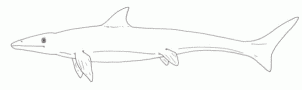
Diversity of Norian Ichthyosaurs: Shonisaurus sikanniensis (above) - length: 23 meters - a gigantic, whale-sized species "Shastasaurus" neoscapularis (Metashastasaurus of Nicholls and Manabe 2001, Callawayia Maisch and Matzke, 2000) (top left) - a close relative of Shonisaurus, length: 3 meters (both Shastasauria) Hudsonelpidia brevirostris McGowan, 1995 (bottom left) a small, advanced, dwarf species, only 1 meter long (Parvipelvia) Illustrations are not to scale illustrations © 2001 Vince R Ward - Prehistoric Pages |
| |
The great diversity of ichthyosaurs is indicated by fossil remains from the Sulphur Mountain and Pardonet formations of British Columbia. Late Triassic ichthyosaurs seem to have belonged to two distinct clades, the medium sized to gigantic shastasaurs, which were the more common group, and the (as yet still mostly) medium-sized Euichthyosauria, including Californosaurus and Parvipelvia; the latter being the ancestors of some later Rhaetian types that ushered in the age of "classic" Jurassic marine monsters.
There is some disagreement regarding the phylogenetic position of the shastosaurs; Nicholls and Manabe 2001 argue that the Shastasauridae form a monophyletic taxon, while Maisch & Matzke, 2000 present them as a grade - transitional between Middle Triassic and typical Early Jurassic forms - rather than a clade. Regardless of their exact status, these were often spectacular animals.
It is hard to believe that the mighty snow-capped Himalayas were once shallow sea beds where tropical invertebrates flourished, and above which fish swam and giant ichthyosaurs frolicked, but such is the case, and fossils found here indicate the continuity in time and wide (probably global) dispersion in space of the shastosaurian ichthyosaurs (specifically shonisaurs).
The partial skull and skeleton of Himalayasaurus tibetensis Dong, 1972 was collected from Norian marine strata near Mt Everest, at an altitude of some 4800 meters above sea-level [Lucas 2001 p.117]. Himalayasaurus is a clear descendent or close relative of the late Carnian Shonisaurus popularis, which it resembles closely in size and build. This species is distinguished by the cutting-edges on its flattened tooth crowns, which are otherwise unknown among ichthyosaurs. The large size (around 15 meters) and extensive cutting edges of the teeth indicate a diet of other tetrapods, and hence a lifestyle perhaps not unlike that of a modern killer whale [Motani et al 1999]. The 10 meter long Tibetosaurus tingjiensis Young, Lui & Zhang, based on a partial skeleton from the Tingi district of Tibet, is indistinguishable in the published diagnosis, and being from the same stratigraphic unit (Langjiexue Group) in nearby localities, are considered by Lucas 2001 p.118 to be synonyms. According to Motani et al 1999, the similarities between H. tibetensis and S. popularis are so small as to indicate the two forms belong to the same genus, the only clear difference between the two being the shape of the tooth crown. If the two species are congeneric, the name Himalayasaurus Dong, 1972 has priority over Shonisaurus Camp, 1976 (being earlier), and the shonisaur species would have to be renamed Himalayasaurus, although it is hoped this won't happen because the name Shonisaurus is already so well known! Maisch and Matzke, 2000 in contrast consider both Himalayasarus and Tibetosaurus as nomina dubia, while in their paper Nicholls and Manabe 2001 use the term Shonisaurus.
Whatever the exact identity of the Himalayan giants, it would be a pretty safe bet to consider them shastosaurs, and almost certainly Shonisaurs. The existence of very similar animals on opposite ends of Pangea (Shonisaurus is known from the Carnian of California and the Norian of Canada) need not surprise us. The great whales of today have no problem swimming across oceans, and 10 to 25 meter Shonisaurs would likewise have swum the Triassic Panthalassian ocean with ease.
Wherever you were in the late Triassic, if you had to go out into the open sea, at the right time, chances are you would glimpse one of these marvelous reptiles surfacing to breath, with a huge spray of water and foam.
Like the Carnian before it, the Norian age was both a time a change and a time of continuity.
Norian freshwater faunas were little changed from those of the Carnian, including various invertebrates, a diverse selection of fishes, sharks, both chondrostian and primitive neopterygian actinopterygians, coelacanths and lungfish, and temnospondyl amphibians and phytosaurian reptiles. These latter, being at the top of the freshwater food chain, were more susceptible to environmental stress than fish and invertebrates. So we find that among the temnospondyls and phytosaurs a number of Carnian forms have disappeared, such as the metoposaurs and most of the phytosaurs (all belonging to the paraphyletic subfamily (or grade) Rutiodontinae, which experienced a dramatic turn-over, similar to the middle of the late Carnian phytosaur extinction. All Norian phytosaurs are of the subfamily Pseudopalatinae, which evolved from rutiodontines, and like them filled a diverse selection of ecological roles.
Originally it was believed that there was a mass extinction at the end of the Carnian, even a meteor impact. However, it seems that some of the apparent extinctions were due to sampling bias, as continental drift means the moist environments that favour the typically Carnian biota are under-represented because the rich fossil localities now correlate with higher and more arid latitudes [Olsen et al]. In Eastern North America (centra Pangea) at least, Norian terrestrial assemblages differed little from those of the Latest Carnian (late Tuvalian / Adamanian)). Ichnotaxa (footprint taxa) found widely across the Newark Supergroup basins in this period include Brachychirotherium (rauisuchian), Apatopus (phytosaur), Rhynchosauroides (lepidosauromorph or basal archosauromorph) Atreipus (ornithischian dinosaur) and Grallator (coelophysid dinosaur) [Olsen and Huber 1998]. These taxa generally represent family or related generic groupings, rather than individual species.
This is not to say there was no turn-over at all. During the Norian age the fortunes of the many types of Carnian terrestrial herbivores seem to have declined. The common and cosmopolitan stagonolepine aetosaurs and hyperodopedontine rhynchosaurs, and the big lizard-like trilophosaurs (which in contrast were limited to western equatorial Lausasia), seem to have disappeared completely. The large kannemeyeriid dicynodonts are still present in moist equatorial West Pangea, as are the dog-like traversodontids. The large aetosaurs are replaced by small ones. The sauropodomorph and ornithischian dinosaurs and the small stocky lizard-like procolophonids are unaffected, and it seems that the extinctions were insignificant in comparison to the diversity of those that were unaffected. [Hunt, et al]
Unlike the semi-aquatic phytosaurs, terrestrial carnivores have done well. The late Carnian rauisuchian Postosuchus kirkpatricki continues into the early or middle Norian [Lucas 1998, Hunt et al], while the medium-sized predator guild was similarly little affected - herrerasaur dinosaurs are known in the Norian, and ornithosuchid "thecodonts", while unknown at this time, are represented in the Late Norian/Rhaetian Los Colorados of Argentina. sphenosuchid crocodylomorphs continue to flourish.
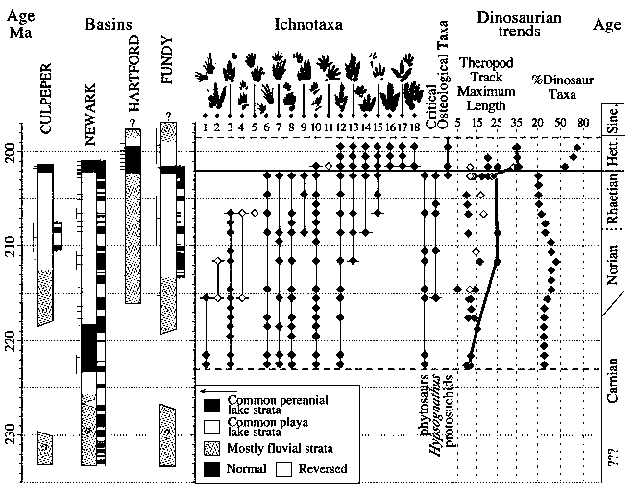
|
Correlation of four key basins of the Newark Supergroup showing the temporal ranges of footprint ichnogenera and key osteological taxa binned into 1-My intervals showing the change in maximum theropod dinosaur footprint length (line drawn through maximum) and percent at each 1-My level of dinosaur taxa. Short, horizontal lines adjacent to stratigraphic sections show the position of assemblages, and the attached vertical lines indicate the uncertainty in stratigraphic position. Solid diamonds indicate samples of footprints, and open diamonds indicate samples with 10 footprints. Horizontal, dashed gray lines indicate the limits of sampling; thick gray line indicates trend in maximum size of theropod tracks; ?, age uncertain. Ichnotaxa are as follows: 1, Rhynchosauroides hyperbates [Lepidosauromorph or primitive archosauromorph]; 2, unnamed dinosaurian genus 1 [Unknown dinosaur, perhaps herrerasaurid]; 3, Atreipus [Ornithischian dinosaur]; 4, Chirotherium lulli [Crurotarsan, possibly aetosaurid]; 5, Procolophonichnium [procolophonid parareptile]; 6, Gwyneddichnium [tanystropheid]; 7, Apatopus [Phytosaur]; 8, Brachychirotherium parvum [Rauisuchian]; 9, new taxon B [Crurotarsan, possibly crocodylomorph]; 10, Rhynchosauroides spp. [lepidosauromorphs]; 11, Ameghinichnus [Advanced synapsid, possibly trithelodontid]; 12, "Grallator" [Small theropod dinosaur]; 13, "Anchisauripus" [Small- to medium-sized, theropod dinosaur]; 14, Batrachopus deweyii [Crocodylomorph]; 15, "Batrachopus" gracilis [Crocodylomorph]; 16, Eubrontes giganteus [Large theropod dinosaur]; 17, Anomoepus scambus [Ornithischian dinosaur]; and 18, Otozoum moodii [Prosauropod dinosaur]. Ma: million years ago Hett.: Hettangian Sine.: Sinemurian M. J. Szajna, B. W. Hartline, in The Great Rift Valleys of Pangea in Eastern North America: Volume 2, Sedimentology and Paleontology, P. M. LeTourneau, P. E. Olsen, Eds. (Columbia Univ. Press, New York, in press). Stratigraphic and magnetostratigraphic columns and correlations are modified from P. E. Olsen, Ann. Rev. Earth Planet. Sci. 25, 337 (1997). Details of vertebrate assemblages among other additional information is provided in the supplemental material available on Science Magazine Online at www.sciencemag.org/cgi/content/full/296/5571/1305/DC1. Correlation with the other rift basin sequences is based on the larger scale magnetic polarity pattern, Milankovitch cycle stratigraphy, palynology, and basalt geochemistry. P. E. Olsen, R. W. Schlische, M. S. Fedosh, Mus. North. Ariz. Bull. 60, 11 (1996). text and graphic © P. E. Olsen, D. V. Kent, H.-D. Sues, C. Koeberl, H. Huber, A. Montanari, E. C. Rainforth, S. J. Fowell, M. J. Szajna, B. W. Hartline, 2002 [ref Olsen et al 2002 [Fig.1] |
The dinosaurs were the big winners of the Norian, increasing in diversity and size. As can be seen from the above diagram, the theropods increase almost four-fold in linear dimensions from the early Tuvalian (mid Carnian) to the Mid Norian, while at least double the number of dinosaurian species are known from any one time span.
The Norian therefore was an age dominated by archosaurs. As we have seen, the larger synapsid herbivores - the lumbering Dicynodonts, and the traversodontid cynodonts - were now reduced in both diversity and range, confined as Carnian relics to a narrow wet tropical band, although still locally common. The medium-sized carnivorous chiniquodontids were replaced by much smaller, and more mammal-like, forms called Dromatheriidae, the ecological equivalent of a shrew. These advanced tiny cynodonts were probably very successful, but remain poorly known, because such tiny remains are only rarely fossilized.
As with the Carnian, the Norian is a long period of time (in fact it is the longest age in the Triassic, in terms of duration) that embraces several distinct tetrapod faunas. We find that a number of different ecological and biogeographic communities existed, developed, and were replaced by new communities, the most dramatic being the sauropodomorph takeover. A review of these is given here. As large predators are relatively rare, we have - following the "empires" of Anderson & Cruikshank 1978, Bakker 1977, etc - defined these by the most common herbivores or (in the case of the sphenodont lepidosaurs or common terrestrial metaposaur Apachesaurus) insectivores. They can be represented as follows:
Description of this first stage is very speculative, due to incomplete strata.
Following the Carnian-Norian turnover, there is an impoverished fauna. Of the large herbivores the rhynchosaurs, trilophosaurs, and stagonolepine aetosaurs are completely extinct, whilst dicynodonts and traversodonts have completely disappeared from the north-west and form eastern Pangea (but still found in in south-west Pangea). Of the carnivores the prestosuchids (represented by the single genus Saurosuchus) are also gone. Among the semi-aquatics , the rutiodontine phytosaurs and the large metoposaurs are extinct, replaced by nicrosaurs and cyclotosaurs (and chigutosaurs in the south). The dinosaurs are the big winners, with several new types appearing.
The West Pangean Aetosaur-Apachesaur Community: As explained, no Earliest Norian terrestrial faunas are known for sure from south-western North America. But it does seem that at some point, perhaps the end of the Carnian, some groups - like the rhynchosaurs and Trilophosaurs on land, and the rutiodontine phytosaurs and large metaposaurs in freshwater, disappeared from this region entirely. In fact, the rhynchosaurs and large metaposaurs seem to have died out worldwide, and no post-Carnian Trilophosaurs are known with certainty. However, a number of latest Carnian (Adamanian) species seem to have continued unchanged right through to the Late Early Norian, so we can assume that they were present at this time, even if fossils are absent. These species would include the large rauisuchian Postosuchus kirkpatricki, the large aetosaurs Desmatosuchus haplocerus and Paratypothorax andressum, the small metoposaurid Apachesaurus gregorii, and small podokesaur dinosaurs (Coelphysis or similar)
The Middle Pangean Metoposaur Community: To the north and east, in Central Laurasia, Metoposaurus stuttgartensis is known from the Lehrbergstuff of Stuttgart-Sonnenberg, Germany. This is the only known Norian metaposaur from Europe. Like Apachesaurus the fragmentary remains belonged to a relatively small animal [Milner 1994, p.11]. It seems then that metoposaurs had been drastically diminished at the end of the Carnian, with only a few specialized forms surviving. Interestingly, in Gondwana, Chigutosaurs take their place (as is evident from late Triassic deposits in India [****), in Europe it is the capitosaurs (Cyclotosaurus) that takes over.
The South-West Pangean Kannemeyeriid-Traversodont Community: In southern Brazil meanwhile, the Catturita Formation of Latest Carnian to Early Norian age, the highest level, informally called the Jachaleria Level [Scherer 1994] includes the kanneymererid Jachaleria candelariensis and the very primitive dinosaur Guaibasaurus candelariensis (which has features indicating affinities with both the sauropodomorph and the Theropoda, implying that it may belong to the ancestral group for both of them; it has been given its own family: the Guaibasauridae. Also found in the Catturita Formation is the earliest known Prosauropod proper, Unaysaurus tolentinoi; preliminary phylogenetic analysis indicates it to be closely related to the European Plateosaurus (Plateosauridae). [Leal et al 2004]
In the arid equatorial west, new advanced phytosaurs (Pseudopalatus) and aetosaurs (Typothorax) appear (although these may have evolved in the preceding stage and not been recorded because of absence of equivalent strata). And for the first time, both herbivorous and carnivorous dinosaurs grow as large as the bigger "thecodonts".
The West Pangean Aetosaur-Apachesaur Community: later Early Norian terrestrial faunas are so far only well known from the middle Chinle Group of south-western North America, where it seems that while some groups - like the rhynchosaurs and Trilophosaurs on land, and the phytosaurs and metaposaurs in freshwater, disappeared entirely, many of latest Carnian species seem to have continued unchanged, alongside a few newcomers. In the Cooper Canyon Formation of Texas we find a transitional Adamanian-Revueltian fauna characterized by the rauisuchians Postosuchus kirkpatricki and Chatterjeea elegans, the large aetosaur Desmatosuchus haplocerus, all late Carnian, as well as a small metoposaurid (perhaps this is Apachesaurus or a relative) and other temnospondyl amphibians, the advanced phytosaur Pseudopalatus buceros, the small ornithischian dinosaur Technosaurus smalli, and the procolophonid Libognathus [Small 2002]
In New Mexico the type Revueltian fossil assemblage of the Bull Canyon Formation in New Mexico is dated Early to Middle Norian, and probably extends over some time, as Hunt 1989 distinguishes two biochrons. The "Anaschisma (here = Apachesaurus?) -Belodon- (=Pseudopalatus) Typothorax-Desmatosuchus-Paratypothorax biochron" includes the rauisuchians Postosuchus kirkpatricki and Chatterjeea elegans, Revueltosaurus (formerly thought to be an ornithithischian dinosaur), a diverse aetosaur fauna consisting of Typothorax coccinarum, Desmatosuchus sp. and Paratypothorax sp., the sphenosuchian Hesperosuchus, and the possible ornithischian dinosaur Lucianosaurus wildi [Hunt 1989, Lucas 1998].
The wide diversity of large aetosaurs suggests that they have taken over the role of big herbivore vacated by the trilophosaurs, rhynchosaurs, and dicynodonts with their disappearance from the region at the end of the Carnian.
A small prosauropod is known from fragmentary postcrania from the Cooper Member of the Dockum Formation of Texas, this was previously thought to be an ornithischian because of the presence of ornithischian teeth (Technosaurus) in this locality. However it seems the two animals were distinct [Hunt and Lucas 1994 p.236]. Neither ornithischians nor prosauropods seem to have been as common or important ecologically as aetosaurs, no more than a single species of ornithischian is known from any one locality, and these dinosaurs are rare throughout the Late Triassic [ibid p.236].
A significant new addition to these faunas is the large (5.5 meter) coelophysid theropod Gojirasaurus quayi, from the late early or middle Norian Cooper Canyon Formation of New Mexico. This was the first of the larger carnivorous dinosaurs, although it is unlikely it ever competed directly with the heavily built Rauisuchians of the time. It correlates with the appearance of larger theropod footprints in the Newark Supergroup during the middle Norian
See also Therrien and Fastovsky 2000 for a description of this and nearby Paleoenvironments (Table 2 in that document includes a list of species)
In this seasonally arid environments, the terrestrial metoposaur Apachesaurus gregorii makes up a common element, while the Carnian aquatic metaposaurs are either very rare or completely absent.
The Middle and East Pangean Aetosaur-Prosauropod-Proterochersid Community: The Lower Stubensandstein fauna of South-west Germany is very similar to that of the Upper Chinle or Upper Dockum of Western North America, and the associated Neshanician stage of the Newark Supergroup of eastern North America (New England, etc). Included here are basic pseudopalatine phytosaurs (Nicrosaurus kampfi), and the large aetosaur Paratypothorax andressum [Rauhut and Hungerbuhler 1998]. These are typical of Revueltian tetrapod assemblages of Lucas 1998. Dinosaurs are represented by the medium- to large-sized (4 to 6 meters long) prosauropod Plateosaurus gracilis, a species which would seem to be directly seem to have been directly ancestral to the later plateosaurs. Temnospondyl amphibians (Cyclotosaurus and Gerrothorax), phytosaurs, and the earliest known turtles (Proterochersis) are all found. No doubt smaller animals were also present. (Aetosaurus ferratus dates from either the Lower or Middle Stubensandstein, the original location is not certain)
No ornithischians have as yet been found, indicating a different community or environment to that of the contemporary West Pangean (SW USA). The only indication of cosmopolitanism is the large aetosaur Paratypothorax andressum which occurs in both regions. Other elements are similar but not identical. There are Cyclotosaur capitosaurs instead of Apachesaur metoposaurids, the phytosaur Nicrosaurus rather than Pseudopalatus, and prosauropods instead of ornithischians. These differences may be the result of geographic isolation between the two faunas.
A similar fauna occurs in southern Gondwana, since the lower part of Dharmaran Formation, India, which might tentatively be dated at around this time, includes Nicrosaurus, a Paratypothorax-like aetosaur, and prosauropods. [Lucas 1998]
The West Pangean Aetosaur-Apachesaur Community: (continued). The upper Bull Canyon Formation in New Mexico represents the Apachesaurus-Pseudopalatus-Typothorax biochron. Filling the medium predator niche is Chatterjeea elegans, which continues from the earlier stage, and a new, small, advanced cynodont, Pseudotriconodon chaterjeei, appears. The ornithischian Lucianosaurus replaces Technosaurus. The upper Petrified Forest Member near San Ysidro might be the same age [Hunt 1989]. The aetosaur Aetosaurus arcuatus (I don't know if this occurs in the upper or lower parts of the formation) is known from both New Mexico and the Newark Supergroup of Eastern North America, indicating that, like its larger cousin Paratypothorax andressorum known from both western North America and Europe (although its long stratigraphic range (late Carnian to middle Norian) means it isn't of much help in determining age of the faunas) Hunt and Lucas 1992, these animals traveled widely cross northern Laurasia
Understanding the nature of vertebrate faunas of south-western North America depends on the interpretation of the stratigraphy. There is disagreement regarding whether the Whitaker (Ghost Ranch) quarry dates from the early Norian or the Rhaetian. If the latter, than the dinosaurs is Eucoelophysis baldwini whose fossil remains are known from the Upper Triassic Petrified Forest Formation of the Chinle Group in north-central New Mexico [Sullivan and Lucas 1999]. This was a small animal closely related to (either a near cousin or perhaps an ancestor) to Coelophysis bauri and the Hettangian Syntarus rhodesiensis. If the former than Eucoelophysis would be Latest Carnian in age. The type horizon of E. baldwini is the upper part of the Petrified Forest Formation, about 45 meters stratigraphically below the locality of the neotype of Coelophysis bauri.
The Central West Pangean Equatorial Aetosaur-Traversodont Community: During the Middle Norian a narrow swath around the Pangean Equator remains home to a persistent Carnian-type biotic province characterized by the presence of small herbivorous traversodont cynodonts, (Plinthogomphodon), rare kannemeyerid dicynodonts, the ubiquitous aetosaur Aetosaurus (Stegomus), a poposaurid rauisuchian similar to Postosuchus, the sphenosuchian crocodylomorph Dromicosuchus, and phytosaurs and temnospondyl amphibians. [Olsen et al]
Elsewhere, fossil footprints indicate the presence of lepidosauromorphs, tanystropheids, Phytosaurs, a possible herrerasaurid and both small and small- to medium-sized podokesaurid theropod dinosaurs [ref Olsen et al 2002].
In places fairly large sauropodomorphs come to dominate the environment (certainly in the Keuper, and it can be expected elsewhere too). The Middle Stubensandstein of Germany, some of the fissure fillings of Bristol, England, the Ørsted Dal Member of the Fleming Fjord Formation, Greenland; the Zorzino Limestone and Forni Dolomite, northern Italy, share very similar faunas (including the same species, or closely related ones) and can probably be considered more or less contemporary.
This stage then sees the appearance of both advanced forms (e.g. Mystriosuchus) and, curiously, a resurgence of several primitive types (Aetosaurus, Proganochelys, and Thecodontosauridae - Efraasia)
The Middle and East Pangean Prosauropod-Proganochelyid-Aetosaur Community. To the east, in Central Laurasia, in the Middle Stubensandstein, which could be considered contemporary with the above, or nearly so, we find a sudden radiation of sauropodomorphs, which increase in both quantitative numbers and diversity. Sellosaurus is now joined by the small, rare Thecodontosaurus and the large (6 meters long) common Efraasia. Curiously both these species are of a more primitive, basal, type, and would appear to be immigrants, as they have no ancestors in the Lower Stubensandstein. Similarly, Proterochersis has disappeared, and is replaced, by a likewise more primitive form, Proganochelys
As for other reptiles, the advanced phytosaur Mystriosuchus now joins the more generalized Nicrosaurus. As with the dinosaurs, this is almost certainly an immigrant (perhaps from the Tethys, where it is also known), as no obvious ancestors are known. These genera were all provincial (however it has been suggested that Nicrosaurus also occurs in North America, although the interpretation of buceros as Nicrosaurus [Zeigler et al 2001] is controversial [see different interpretation in Hungerbühler 2002 where it is considered Pseudopalatus], the aetosaur Aetosaurus ferratus is different in species but generically similar and again closely related to its west Laurasian counterpart. Here we also find the Rauisuchian Teratosaurus can be compared to Postosuchus and Chatterjeea.
Two small theropod dinosaurs are also known from this level, the small Procompsognathus triassicus and the medium-sized Halticosaurus longotarsus, the latter too fragmentary identify properly (nomen dubium).
These early dinosaurs lived alongside amphibians (Cyclotosaurus posthumus continues from the Lower Stubensandstein), aetosaurs, and new species of phytosaurs (Mystriosuchus) and sphenosuchids (Saltoposuchus).
This European (Central Laurasian) fauna extended as far as East Laurasia (Thailand), where one finds in the Huai Hin Lat Formation (dated as Norian on the basis of plants and ostracods [deBroin et al 1982) Actinopterygii, lungfish, the temnospondyl Cyclotosaurus robustus, indeterminate plagiosaurs, a distinct species of Proganochelys (P. ruchae) , and phytosaurs Mystriosuchus and "a Beledon [=Nicrosaurus) -like form" [Buffetaut et al 2000]
Marine and Terrestrial Reptiles of the Late-Middle Norian Tethys (Zorzino Limestone) |
 |
Center-left (crawling into the water) is the placodont Psephoderma alpinum, behind which is the phytosaur Mystriosuchus planirostris, which also occurs in the Middle Stubenstein, while in the middle forground the thalattosaur Endennasaurus acutírostrís pokes its head out of the water. In the left foreground is the insectivorous prolacertiform Langobardisaurus pandolfii, while in the right foreground the sphenodontid Diphydontosaurus stalks a beetle. In the middle background is the aetosaur Aetosaurus ferratus (known from this locality only from a small fragment of dorsal armour). To the right are the arboreal archosauromorphs Drepanosaurus and Megalancosaurus. Above fly several pterosaurs. illustration © copyright Dr Silvio Renesto original url
|
The Tethyan Pterosaur-Drepanosaur Community (above) When we move south to the Tethyan coast, and west to western Laurasia, we find another unique ecosystem. A distinct endemic fauna has been found at the Zorzino Limestone and Forni Dolomite in northern Italy, and represents a tropical coastal environment. Being marine sediments, these can be very accurately dated by ammonoid and conodonts. The Zorzino Limestone turns out to be very close in age to the Alaunian-Sevatian boundary, being placed in the younger part of the Mesohimavatites columbianus zone. The Forni Dolomite, also Mesohimavatites columbianus zone, is dated at Middle-Late Alaunian; it contains different species of Pterosaurs and so may be slightly older. The common presence of the aetosaur Aetosaurus ferratus and the phytosaur Mystriosuchus planirostris indicates that the Middle Stubensandstein and the Zorzino Limestone are probably the same age - Latest Middle Norian - or at least fairly contemporary.
The Thecodontosaur Sphenodont Community During the late Triassic and early Jurassic in middle north Pangea, Carboniferous limestone caves and crevices in presumably upland regions, served as traps into which animals would stumble into, fall, and be unable to get out. Eventually their bodies were mummified, preserved, and fossilized as the caves filled with sediments. These became the Anglo-Welsh (and French and German) limestone fissure fillings, which have provided fossil remains of a diverse range of small reptiles and early mammals. Although traditionally dated as "Rhaetic" or "Rhaeto-Liassic", it is now considered that many of these faunas are actually Middle or Late Norian, and some may be as early as the late Carnian onwards. In these regions away from the broad floodplains and braided streams that favoured the large sauropodomorphs, a provincial Thecodontosaur community developed, perhaps as a subset of a larger Prosauropod community. Its endemic nature makes this fauna difficult to correlate with that of larger middle and east Laurasia. Representative forms include Thecodontosaur sauropodomorphs, Sphenosuchid (Terrestrisuchus) crocodylomorphs, gliding lepidosaurs (Keuhnosaurus), sphenodonts, advanced cynodonts, early mammals, and other small animals. Comparisons are sometimes made between Terrestrisuchus and Thecodontosaurus and the Middle Stubensandstein Saltoposuchus and Thecodontosaurus, in order to suggest a similar date for the Bristol fissure fillings, but similarity does not guarantee they are contemporary; Terrestrisuchus is now considered distinct from Saltoposuchus [Fraser et al 2002], and the Stubensandstein "Thecodontosaurus" is too fragmentary to identify with that genus with certainty.