The Norian Age - 3
Tetrapods of the early Late Norian (Sevatian) - Age of Sauropodomorphs
During this period (Late Norian to Rhaetian) large sauropodomorphs dominate most terrestrial environments (except the north-west). Often a single taxon will be very common: Plateosaurus in Europe / Central and North Laurasia, Euskelosaurus in Southern Africa / South Gondwana, and Riojasaurus in Argentina / South-West Gondwana. This is not to say that a number of other taxa are not often present as well, but these are always much fewer in number. Earlier types of aetosaurs and phytosaurs die out, while new phytosaurs, aetosaurs, and dinosaurs (Syntarsus) appear.
The West Pangean Aetosaur-Apachesaur Community: (continued). Animals of west equatorial Pangea seem to be best represented by the diverse remains discovered at the Whitaker (Ghost Ranch) quarry (Rock Point Member of the Chinle Formation, New Mexico), although a little to the north the Chinle Church Rock Member (Utah) might be considered a similar age [Benton 1994].
This can be considered then the earliest date of the Apachean Land Vertebrate Age, as indicated by the presence of the phytosaur Redondasaurus Lucas 1998]. Curiously, the predominance of large sauropodomorphs that characterizes the rest of the world is absent here, this indicates either locally distinct conditions or an incorrect date for these strata.
The Low Latitude Central-West Pangea Hypsognathus Community. Further east, and north of the high rainfall band, in what is now eastern USA (Newark Supergroup), the Laurasian desert continues, but here instead of the typical plateosaur community is a more restricted assemblage characterized by the procolophonid Hypsognathus [Lucas 1998], although fossil footprints indicate the presence of lepidosauromorphs, tanystropheids, Phytosaurs, Rauisuchians, crocodylomorph-like crurotarsans, advanced Crocodylomorphs, Ornithischian dinosaurs, and both small and small- to medium-sized podokesaurid theropod dinosaurs [ref Olsen et al 2002]. Curiously, ornithischians rather than sauropodomorphs seem to be the main herbivore here.
In Europe the Late (Upper) Stubensandstein witnessed a further increase in dinosaurs. Plateosaurus is ubiquitous, having descended from and replaced the earlier forms. It is represented by a single species, Plateosaurus longiceps (P. trossingensis is a junior synonym [Galton 2001, Galton 2002]) and accompanied by phytosaurs and the long-lived turtle Proganochelys quendstedti, which continues from the Early and Middle Stubensandstein. We can assume many other animals were also present, although they have not been preserved as fossils.
According to Dr Bob Bakker, the pattern of emergence of the anchisaur-plateosaur empire follows very closely that of the earlier lystrosaurid-kannemeyerid empire. As Bakker explains, in the faunal zone following the Carnian extinctions (13 in Fig. 2) prosauropod dinosaurs of several closely related families (T and U in Fig. 2) make up nearly all big herbivore specimens.
"In any one local basin, the diversity appears to be very low, reminiscent of that of the Lystrosaurus Zone; usually one genus of big prosauropod dominates the collections, although various growth stages sometimes have been recognized as distinct genera (Rozdestvenski). Surprisingly, the top predators of these early prosauropod zones are holdovers from the mid-Triassic - ornithosuchid and rauisuchid thecodonts (H and L in Fig. 2). Advanced theropod dinosaurs take over this trophic role at the Triassic-Jurassic boundary"
This ecological community probably extended for some distance, since prosauropods and turtles are common in the Fleming Fjord Formation (Malmros Klint and Ørsted Dal Members) of East Greenland, where a diverse assemblage of fossil vertebrates very similar to that of the middle Norian Stubensandstein of Germany has been found [ Jenkins et al 1995, Lucas 1998 ] including fish, temnospondyl amphibians (Gerrothorax, Cyclotosaurus), turtles (c.f. Proganochelys sp.), sphendontids, aetosaurs (Aetosaurus ferratus, Paratypothorax andressi), a pterosaur, prosauropod (Plateosaurus) a theropod dinosaur and footprints, and several species of mammals (Kuehneotherium, c.f. Brachyzostrodon, and Haramayavia) . During the Late Triassic, Greenland and continental Europe were adjacent during time, and dinosaurs and other animals clearly migrated widely. The environment was controlled by Milankovitch cycles, with climatic conditions varying from humid to seasonal to arid. Because a number of animals of different time periods are represented, it is likely that several stratigraphic levels are represented here.
Tetrapods of the late Late Norian (Sevatian) - Age of Sauropodomorphs
The Middle Pangean Prosauropod-Proganochelyid (Plateosaur) Community In middle Laurasia (middle latitude north-central Pangea) the formerly mixed and diverse fauna becomes increasingly dominated by sauropodomorph dinosaurs at the end of the Norian. This appears to be a global trend. The Plateosaur community extended widely throughout all of central Laurasia, as the most common plateosaur species (Plateosaurus longiceps, not P. engelhardti as usually identified - see notes on Plateosaurus, below) is known from Germany, France, Switzerland and Greenland. At this time, the climate in Greenland was very lush, and a rich assortment of plant species thrived (some 200 species are known from the Scoresby Sound area of eastern Greenland, although this represents a stratigraphic sequence extended in time, so not all were contemporary) in the warm and wet (even though high latitude (45 - 50 degrees north) climate. Yet the genus Plateosaurus is found in this part of the world, from the Fleming Fjord Formation, just as in the deserts, and as we have seen is associated with typical Keuper species.
In Germany, the Knollenmergel is almost entirely dominated by Plateosaurs. Three species are known - the common Plateosaurus longiceps, the rare Plateosaurus engelhardti (known from Bavaria, probably a geographically distinct form), and a new genus and species Ruehleia bedheimensis [Galton 2001, Galton 2001b, Galton 2002] which however appears to be very similar to Plateosaurus [Headden 2003]. A fourth species of plateosaur, P. erlenbergiensis from this time is known from incomplete remains and hence indeterminate, it is probably a synonym of one of the others. But regardless of how many species one counts, there is little variation, and only one dominate species.
These large dinosaurs are accompanied by phytosaurs - Angistorhinopsis ruetimeyeri replaces the earlier Mystriosuchus and Nicrosaurus - and the ubiquitous early turtle Proganochelys. Cyclotosaurus robustus has been replaced by Cyclotosaurus carinides, and the palgiosaur Gerrothorax pulcherrimus by a related species, Plagiosaurus depressus
Thus we find in middle Laurasia (middle latitude north-central Pangea) the formerly mixed and diverse fauna becomes increasingly dominated by sauropodomorph dinosaurs at the end of the Norian. This appears to be a global trend. The Plateosaur community extended widely throughout all of central Laurasia, as the most common plateosaur species (Plateosaurus longiceps, not P. engelhardti as usually identified - see notes on Plateosaurus, below) is known from Germany, France, Switzerland and Greenland. At this time, the climate in Greenland was very lush, and a rich assortment of plant species thrived (some 200 species are known from the Scoresby Sound area of eastern Greenland, although this represents a stratigraphic sequence extended in time, so not all were contemporary) in the warm and wet (even though high latitude (45 - 50 degrees north) climate. Yet the genus Plateosaurus is found in this part of the world, from the Fleming Fjord Formation, just as in the deserts, and as we have seen is associated with typical Keuper species.
In the trees and the underbrush living alongside these large dinosaurs were many smaller animals. The locality of Saint-Nicolas-de-Port (Late Norian to Early Rhaetian in age) is famous for having yielded numerous mammal teeth.
The South Pangean Melanorosaur-Sauropod (Euskelosaurus) Community: (High Latitude South Pangea) Going south to Gondwana, the Lower Elliott Formation of Southern Africa provides a glimpse of a rich assemblage of advanced early dinosaurs. The Lower Elliot is traditionally dated as Late Carnian on the basis of Dicrodium plants and the presence of typically Carnian elements like Traversodonts. However the advanced sauropodomorph elements indicate a later date [Lucas and Hancox 2000], and there is no reason why the dicrodium biota could not have continued in Gondwana well into the Norian.
In the Lower Elliott of Southern Africa (south Gondwana) then, we find a high latitude location with at least four different Proto-Sauropod and basal Sauropod genera (Melanorosaurus, Euskelosaurus, Blikanosaurus, and Antetonitrus). There are no mastodontosaurid (Cyclotosaurus) or plagiosaur amphibians; instead, brachiopods fill the role of medium to large semi-aquatic predator. Intriguingly, and adding to the distinct Gondwanan flavour of this community, large synapsid herbivores (a kannemeyerid dicynodont (represented by footprints) and the large cynodont Scalenodontoides) are also present, although rarer than the dinosaurs. The dominance of giant dinosaur herbivores (in this case Euskelosaurus is especially common) shows an ecological equivalence with the Plateosaur community to the north, and we can assume that these large animals were found worldwide. Again, the climate is seasonal monsoonal (savannah-like) to semi-arid. One also finds in Gondwana large rauisuchians - represented by Fasolosuchus in the west (Argentina) and "Basutodon" further south and east. Podokesaurid (= Coelophysid) theropods are indicated by the presence of numerous small to medium-large sized tridactyl footprints (Grallator and larger forms, the nomenclature of the latter (by Ellenberger 1970) is very oversplit.)
Also in Gondwana, the slightly younger (probably Rhaetian) Los Colorados of Argentina reveals a similar preponderance of large sauropodomorphs. It would seem likely in any case that the Sauropodomorph fauna continues through to the Rhaetian with little change; although the extreme provinciality of this time makes comparative dating difficult (so some formations attributed to Late Norian may be Rhaetian, and vice versa).
A Norian Bestiary
|
|
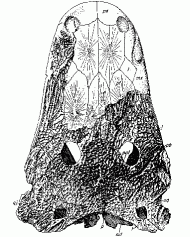
Cyclotosaurus posthumus - skull length 53 cm
Stubensandstein (middle Keuper) of Pfaffenhofen, Bavaria
|
Cyclotosaurus sp.
|
With the drying of the climate across the Euramerican landmass, due to continental movement, the Metoposaurs die out, apart from the small terrestrial Apachesaurus of the western part of the northern Laurasia province. The Cyclotosaurs (family Capitosauridae or Mastodonsauridae) are more successful and continue to flourish in Europe. There the large Cyclotosaurus robustus of the Carnian has been replaced by the related Cyclotosaurus posthumus. It differs from its predecessor in certain minor details, like a more narrower and more coarsely sculptured skull, a slightly narrower snout, and more lateral (sideways) nostrils [Welles and Cosgriff, 1965, p.45]. Cyclotosaurus posthumus is known from the Stubensandstein of Germany, the Fleming Fjord Formation of Greenland, and the Huai Hin Lat Formation of Thailand. It was a large animal, with a skull length of over half a meter, and an overall length of around two to two and a half meters in length. These animals probably lay quietly on the bottom of ponds and shallow rivers, snaring unwary passing fish and small aquatic tetrapods with a sudden opening of the huge mouth. While adults were probably large enough to be safe, the young would have fallen prey to the phytosaurs that frequented the same waterways.
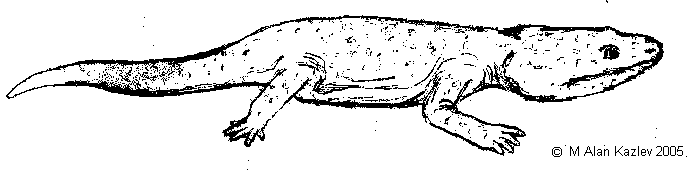 |
Apachesaurus gregorii
|
Most Metoposaurs were large aquatic animals with large heads and small weak limbs. A prominent exception was the diminutive form (skull 25 cm or less) Apachesaurus gregorii (Hunt and Santucci 1993, Hunt 1998), known from abundant intercentra, several partial skeletons, partial skulls, clavicles and interclavicles found in the Chinle Formation of Arizona and New Mexico and the Dockum Group of Texas. Apachesaurus had a much more terrestrial habit than other members of its family, comparable perhaps to the Permian dissorophids. This animal is rare in the late Carnian (Adamanian), where large metoposaurs predominate, but is very common in the Norian of the American southwest [Milner 1994]. It is the most common element of the Painted Desert Member fauna (Chinle Formation). It appears to have been geographically restricted to that region, as it has not been found outside what was then western equatorial Pangea. It seems to have filled the ecological role of semi-aquatic insectivore/small carnivore.
 |
|
Gerrothorax pulcherrimus, from the Stubenstein (Middle Norian) of Stuttgart, SW Germany; Fleming Fjord Formation of Greenland
|
The advanced temnospondyls Plagiosaurus and Gerrothorax were medium-large (around a meter long) short-headed amphibians of the family Plagiosauridae that lived alongside Cyclotosaurs and phytosaurs. They are known not only from the Ladinian to Rhaetian of Europe and (Norian) Greenland, but also the Huai Hin Lat Formation (Norian) of Thailand, and so can be assumed to have had a wide distribution throughout northern middle to north-east Pangea. Plagiosaurs were even better adapted to a totally aquatic existence than the cyclotosaurs; in fact the presence of gills means they were unable to live out of water at all. Gerrothorax fed on fish, invertebrates, and smaller amphibians and reptiles, while themselves falling prey to the phytosaurs with which they shared their environment. Indeed, considering that they seem to have had no natural defenses against phytosaurs, these large Temnospondyls did remarkably well throughout the entire period of the Late Triassic.
 |
|
|
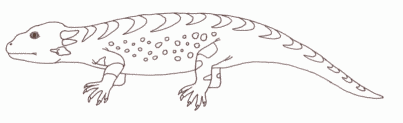
Libognathus sheddi is a representative of the advanced procolophonids; small, squat lizard-like reptiles which flourished in arid (desert or semi-desert) environments of the late Triassic [Olsen et al], and the first procolophonid to be described from the Late Triassic of the southwestern United States. Libognathus' remains were found in the late Early Norian of Cooper Canyon Formation (Dockum Group) near Post, Texas. It is one of a number of North American procolophonids represented solely by tooth-bearing jaws [Modesto S.P. ; Damiani R.J. ; Sues H. , 2002] These have provided an understanding of tooth replacement and implantation in procolophonids. The teeth set in sockets and firmly anchored by spongy hone of attachment (this is known as the protothecodont condition), and are replaced in a typical reptilian pattern. Libognathus is closely related to the Late Norian and Rhaetian Hypsognathus [Small 1997]
The First Turtles
 |
Illustration from Life Before Man by Zdenek V. Spinar, illustrated by Zdenek Burian |
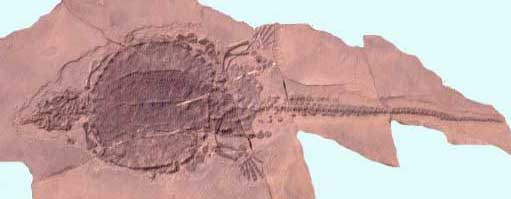
The Norian age was remarkable for the appearance of the Chelonia (turtles) which seem to have emerged out of nowhere. In the Lower and Middle Stubenstein of the German Keuper, they make up a large proportion of local fauna [Benton 1986]. Two species are known. Proganochelys quendstedti is the more common and well known, and a representative of the most primitive lineage of turtles (family Proganochelyidae). It is a largish chelonid, the overall animal being up to about a meter. The skull, although typically chelonid in its robust and solid construction, toothless jaws, horny beak, and other features, still retains a few primitive characteristics not found in more advanced forms, such as tiny teeth on the palate (roof of the mouth). The broad domed shell differs from that of later turtles only in the presence of small marginal spiky plates that projected outwards, and the openings being too small to withdraw the neck and legs into. Instead, the limbs were protected by the spiked plates, and the neck by spiny scales. This long-lived species, which has in the past also been known under the names Psammochelys, Stegochelys, and Triassochelys, continued to the end of the Norian (Knollenmergel). [Gaffney 1986] A related form is known from the Fleming Fjord Formation of Greenland [Lucas 1998]; it would not be surprising if this is the same species. Proganochelys ruchae is a contemporary (Norian) species from Huai Hin Lat Formation of Thailand illustrates the wide (possibly worldwide) distribution of this genus.
Despite appearing even earlier than Proganochelys, Proterochersis robusta is a more advanced form, the earliest known of the side-necked turtles or pleurodires. Unlike Proganochelys the lower shell or plastron has large openings to allow the limbs to retract (although the ability to retract the head and neck inside its shell did not develop, curiously, until the Cretaceous).
The presence of two very different types of turtles even at this time, and the implication of a third from the ghost-lineage formed by an intermediate Rhaetian form Palaeochersis, as well as the primitive Hettangian (early Jurassic) Australochelys, shows that the Chelonia had undergone an extensive evolutionary radiation in the early Norian, if not before. One might hazard a guess that their sudden appearance and abundance in the Norian was due to the disappearance of rivals or predators in the Carnian-Norian turnover. These early turtles clearly lived in a terrestrial environment [Joyce and Gauthier 2004], and it was only later in the Jurassic that the group moved to a semi- and fully-aquatic lifestyles.
Norian Chelonida
Proganochelys quendstedti
Middle and Upper Stubenstein and Knollenmergel; Fleming Fjord Formation
late Middle to Late Norian
Proganochelys ruchae deBroin 1984
Proterochersis robusta Fraas 1913
maximum carapace length of 33 cm
Giant Ichthyosaurs
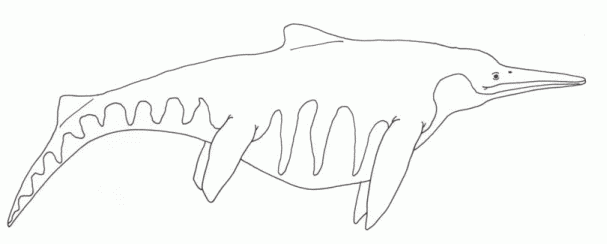 |
Shonisaurus, the lord of the late Triassic seas.
Length: 10 to 25 (or more) meters
Known stratigraphic range: Late Carnian to Norian
Distribution: Panthalassian (world-wide).
|
In a dramatic scene from an episode of Walking with Dinosaurs, "the Cruel Sea", viewers saw a gigantic pliosaur swallow a theropod dinosaur. According to the WWD team, Liopleurodon, the giant pliosaur in question, was 25 meters long, the largest carnivore ever to live.
The truth is more disappointing. No pliosaur is known to have exceeded half that length, and 7 to 8 meters (the size of a modern adult killer whale) was standard. This would seem to be the optimal size of giant predatory marine animals.
However, during the late Triassic, there lived an animal that really was as big as the WWD pliosaur. This was Shonisaurus sikanniensis, whose gigantic remains is known from the Middle Norian Pardonet Formation, northeastern British Columbia. [Nicholls and Manabe 2004]
As well as being a lot bigger, S. sikanniensis differs from S. popularis in the shape of the scapula (shoulder bone) and relatively longer vertebral centra (indicating perhaps a more elongate body). Study of this new species, along with re-examination of S. popularis, indicates that Shonisaurus was not as deep-bodied as previously reconstructed. The teeth seem to have been present only in juveniles. The tendency towards toothless is found in some Jurassic ichthyosaurs as well.
Shonisaurus sikanniensis is the largest marine reptile known, and the largest forms are for that matter the largest type of reptile known (exceeding the biggest terrestrial sauropods in size), the largest carnivorous animal, and among the largest animals ever to live. It is considerably larger than the late Carnian Shonisaurus popularis, the previous record holder, perhaps an ancestor or close relative. A specimen collected by the Tyrrell Museum in 2000 is 21 meters long, and, incredibly, even larger specimens are known; putting these animals in the size range of the larger baleen whales [Nicholls 2003]. But unlike baleen whales, which are filter feeders, Shonisaurus was a carnivore, and very probably preyed on smaller (but still large) vertebrates, although it is not impossible that, like modern sperm whales, they may also have fed on giant cephalopods, assuming such existed at the time (coleoidea evolved in the Devonian, giant ammonites are known from the Cretaceous). The skull is massive, and the eye sockets alone are well over a meter across [Headden, 2004]. In life, had the largest eyes of any animal that ever lived.
The sunken carcasses of these giant animals supported whale bone-like chemosymbiotic communities, and provided nutrients for representatives of several different types of bivalves, including the Lucinidae and Thyasiridae, which are represented in modern and fossil cold seeps and in association with sunken carcasses, and Halobia and Monotis, which probably harboured chemosymbiotic bacteria [Johnston et al 2000]
These huge ichthyosaurs were part of a diverse ecosystem, with at least five genera of ichthyosaurs represented in the Pardonet formation, the two most common being Shonisaurus and the related Callawayia [Nicholls 2003].
These gigantic animals seem to have became extinct at the end of the Norian, and were replaced by other, smaller, genera.
Shonisaurus tibetensis Dong, 1972
body length from 10 to 15 meters
body weight up to 40 tonnes
= Himalayasaurus tibetensis; includes Tibetosaurus tingjiensis [see Lucas 2001 p.117]
Shonisaurus sikanniensis Nicholls and Manabe 2004
northeastern British Columbia.
Marine predator/super-predator
body length up to 21 meters and more
body weight up to ~100 tonnes and more
the largest known marine reptile
Callawayia neoscapularis Maisch and Matzke, 2000
northeastern British Columbia.
body length up to 3 meters
Placodonts
Psephoderma alpinum, is better known from the Rhaetian, but has also more recently been found in the late Middle Norian deposits. Like many of the Rhaetian forms, this early individuals lived in near-shore Tethys waters. However Norian specimens are at least 50% longer in linear dimensions than Rhaetic forms, so it is possible that these might be a different subspecies or even species. Psephoderma was a sort of reptilian ray, with a broad, flat body protected by a discoidal carapace. As with many other placodonts, the teeth are adapted to crushing shellfish, and it probably fed on mussels like Isognornon and Modiolus.
Psephoderma alpinum (large)
Ray-analogue - flattened bottom-dweller that fed on shellfish.
Largest complete specimen is 180 cm long, but isolated teeth indicate it grew larger
Lepidosauromorphs
Kuehneosaurus is the epynomous representative of the Triassic gliding lepidosaurs or Kuehneosaurs. Although usually described as a lizard, and closely related, it nevertheless belongs to an earlier branch of the Lepidosauria, a lizard uncle rather than a true lizard, being equally related to both the Sphenodonta and the Squamata. Like its close relative the late Carnian Icarasaurus, it glided from tree to tree using wings made from skin stretched over elongated ribs. This was the same adaptation acquired by the Permian basal diapsid Coelurosauravus and the recent gliding lizard flying dragon Draco volans. The fact that three groups of small reptiles independently evolved the same characteristic shows the successful nature of this specialized adaptation
Pant-y-ffynnon fissure filling
Diphydontosaurus avonis and Planocephalosaurus robinsonae were very similar, very primitive Sphenodonta known from the Middle or Late Norian and Rhaetian of England and Wales (fissure fillings). Sphenodonts were both common and diverse during the Morian, and a number of species are known. These small animals were the Triassic equivalent of lizards; and in fact true lizards (Squamata) did not appear until the Middle Jurassic. There small size makes preservation difficult, and added to that there is "collector bias" in that large and obvious fossils are recovered more easily in favour of smaller and more modest (and often tiny and difficult to notice) remains. It is only in exceptional or carefully screened deposits that they are recovered.
Diphydontosaurus and Planocephalosaurus both belong to the paraphyletic family Gephyrosauridae, and constitute the most primitive grade of Sphenodonta
A juvenile Diphydontosaurus (illustrated above) belonging to a different species to D. avonis is also known from the latest-middle Norian of Tethyean Northern Italy, and is the only lepidosaur found in that locality. It seems that the Tethyan ecomorph was dominated by archosauromorphs, whereas the presumably drier inland and upland environments were frequented instead by Sphenodonts.
Archosauromorphs
 |
|
|
Among the more peculiar of the Norian lizard-like archosauromorphs (prolacertiforms) were the Drepanosauridae, a clade of reptiles that include the Tethyan Megalancosaurus and Drepanosaurus (known from the Dolomia di Forni and Calcari di Zorzino of Northern Italy), the west Pangean/Laurasian Dolabrosaurus (Dockum Formation) and Hypuronector (Lockatong Formation of New Jersey), and a number of other forms. Originally these animals were interpreted as aquatic swimmers because of the deep tail, but it is much more likely they were arboreal; the tail is laterally stiff and dorsoventrally mobile at its base. [Renesto, 2001]. Megalancosaurus is perhaps the best known form. It was about 30 cm in length, possessed long sharp hooked claws, opposable digits and a chameleon-like prehensile tail. The triangular, bird-like skull is set on a ling neck, and there is a sort of hump for muscle attachment at the shoulders. All of which gave this creature a rather strange appearance. Drepanosaurus was a bit larger and differed in some details; the skull is not known but the skeleton has parallels with the arboreal pigmy anteater, it is likely that Drepanosaurus used its large claws to break open bark to get at insects. Being small and living in an environment were preservation depending on their falling into water and being quickly buried in anoxic silt, these little diapsids were probably a lot more common than their few fossil remains indicate.
Links:
Megalancosaurus - Dr Silvio Renesto
Megalancosaurus preonensis - Dan Bensen
Continued on Next Page
page uploaded 10 May 2005
text content by
M.Alan Kazlev 2005, Creative Commons Attribution Non-Commercial License versions
1.0 and
2.0
images on this page are copyright respective artists or authors