| Palaeos: | Demospongiae | |
| Porifera | Demospongiae - 2 |
| Page Back | Unit Back | Metazoa | Metazoa Dendrogram | Metazoa References | Pieces | Taxon Index |
| Page Next | Unit Next | Unit Home | Unit Dendrogram | Unit References | Glossary | Time |
PORIFERA |--Archaeocyatha `--+--Stromatoporoidea `--+--Calcarea `--+--Hexactinellida `--+--Demospongiae | |--Tetractinomorpha | `--Ceractinomorpha `--+--Homoscleromorpha `--EUMETAZOA |
Introduction Spicules: a New Look Sponge Skin Rootlets and Relationships New Developments History The Sequence of Sponges
|
 Special
Credit: Special thanks to Profs. Sally Leys and Jean-Yves Exposito for
answering questions without knowing how the information would be misused.
Special
Credit: Special thanks to Profs. Sally Leys and Jean-Yves Exposito for
answering questions without knowing how the information would be misused.
This section is actually about demosponge collagens. However, having adopted this alliterative title, we feel compelled to say something about sponge "epithelium" in general. As we have mentioned elsewhere, the term "epithelium" is placed in quotation marks because sponge skin generally has no basement membrane. It seems that, absent a basement membrane, quotation marks are needed to hold the tissue together.
Homoscleromorpha have Type IV collagen. What ought to be holding the tissue together is type IV collagen. Every non-sponge animal has type IV collagens and basement membranes. However, among sponges, only the Homoscleromorpha (Oscarella and that crowd) have basement membranes and type IV collagen. Boute et al. (1996); Exposito et al. (2002). This is a very useful and important fact. It demonstrates that the homoscleromorphs are indeed very advanced sponges. The usual moral drawn from this data is that the homoscleromorphs are thus not demosponges as once thought, but more closely related to Calcarea -- consistent with the latest molecular sequence phylogenies. That is rubbish ... but perhaps we are being too subtle. It is not only wrong, but absolutely and perversely wrong.
Type IV collagen is similar to Spongin. Type IV collagens are indeed special -- and quite distinct from the fibrillar collagens found in all animals, sponge and non-sponge alike. On the other hand, type IV collagens are quite similar to spongin, a collagen thus far found only in demosponges. So, the proper conclusion is that homoscleromorphs are close relatives of demosponges, just as they appear by morphology, and that the collagen data is egregiously inconsistent with sequence phylogenies. Of course phrases like "quite similar" are mushy and often abused. However, any (hypothetical) readers can examine, for example, the review by Exposito et al. (2002) at this direct link and make up their own minds.
Recently, this dissonance has become even worse. It turns out that spongins
are not merely homologous to type IV
 collagens. Rather spongin is
itself conserved in Metazoa, as the basal family of a large class of short-chain
collagens widely distributed in animals.
Aouacheria
et al. (2006). These short-chain collagens had escaped notice until
recently because they are absent in the two largest and most-studied clades,
Vertebrata and Ecdysozoa.
collagens. Rather spongin is
itself conserved in Metazoa, as the basal family of a large class of short-chain
collagens widely distributed in animals.
Aouacheria
et al. (2006). These short-chain collagens had escaped notice until
recently because they are absent in the two largest and most-studied clades,
Vertebrata and Ecdysozoa.
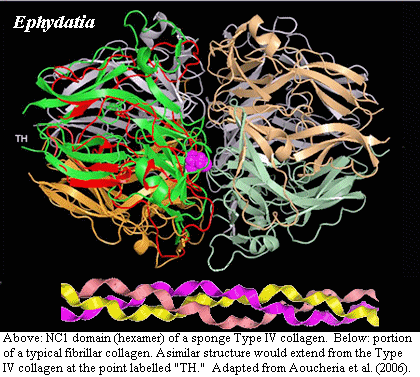
Type IV collagen is quite distinct from fibrillar collagens. It is worth some emphasis that type IV and short-chain collagens are so different from fibrillar collagens that they are scarcely collagens at all. The characteristic triple-helical domain of collagen, with its repeating GPP* (where P* = hydroxyproline) motif, seems to have been grafted onto one end of a very different protein which forms a series of folded β-sheets, tightly interconnected by cystine disulfide bridges at moderately well-conserved positions (the NC1 domain). This non-collagenous domain has itself undergone duplication (NC1a and NC1b), further diluting the triple helical portion. The triple helical portion is also interrupted and terminated by two short additional non-collagen domains (NC2 and NC3, respectively) which likewise link to other elements via cystine disulfides. Aouacheria et al. (2006). In fact, some of these short chain species have no triple helical domain at all. Id. (see their Figure 1).
Finally, just to kick this dead horse one more time, Aouacheria et al. believe that the short-chain "spongin-like" collagen from at least one demosponge, Ephydatia, is in fact better described as a Type IV collagen. If so, the Homoscleromorpha no longer have a monopoly on these basement membrane collagens. Probable basement membrane layers have also been identified in larval demosponges and there are rumblings that this may be a character more widely distributed in the Demospongiae. Eerkes-Medrano & Leys (2006).
In the interests of full disclosure, we should mention that things are less clear than we would like. For example, it would increase our confidence a good bit if some homoscleromorph species made both type IV collagen and spongin. That may be the case, and the species is simply known as a demosponge, as were all homoscleromorphs until not long ago. On the other hand, nothing about sponge phylogeny is easy, so why should the collagen part be any different?
Homoscleromorphs are probably demosponges. The correct lesson to be drawn from this material is that homoscleromorphs are probably demosponges after all, or represent a closely allied clade. It seems likely that the molecular phylogenies are correct in placing Homoscleromorpha as the living sister group to Eumetazoa. Although, as we will briefly mention later, there is one other contender for this spot -- also a demosponge clade. The molecular sequence work cannot be reconciled with the collagen results, which appear far too tidy and far too complex to be the work of chance. But we have a number of issues to cover before drawing further conclusions.
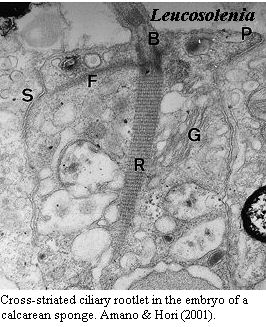
 Cross-striated
ciliary rootlets are the only morphological character joining Calcarea,
Homoscleromorpha and Eumetazoa. One
of the many embarrassments faced by the sponge molecular sequence phylogenies is
the paucity of plausible morphological synapomorphies uniting
Calcarea and
Homoscleromorpha. The thought seems to be that one sponge is pretty
much like another, so homoplasy is probably rampant. That seems a counsel
of desperation -- possibly correct, but we have not yet reached that level
of frustration. However, the supposed clade Calcarea +
Homoscleromorpha + Metazoa does have at least one arguable synapomorphy.
Sperling et al.
(2006). The
rootlets of at least some cilia are banded, or in micro-speak, "cross-striated"
with a periodicity of 40-80 nm. No such cross-striations are found in
Demospongiae [12].
We don't mean to be too flippant about this issue. It is a serious problem
for our hypothesis, and one for which we lack a complete answer. On the
other hand, it isn't all that compelling an argument for a
Calcarea-Homoscleromorpha connection, either.
Cross-striated
ciliary rootlets are the only morphological character joining Calcarea,
Homoscleromorpha and Eumetazoa. One
of the many embarrassments faced by the sponge molecular sequence phylogenies is
the paucity of plausible morphological synapomorphies uniting
Calcarea and
Homoscleromorpha. The thought seems to be that one sponge is pretty
much like another, so homoplasy is probably rampant. That seems a counsel
of desperation -- possibly correct, but we have not yet reached that level
of frustration. However, the supposed clade Calcarea +
Homoscleromorpha + Metazoa does have at least one arguable synapomorphy.
Sperling et al.
(2006). The
rootlets of at least some cilia are banded, or in micro-speak, "cross-striated"
with a periodicity of 40-80 nm. No such cross-striations are found in
Demospongiae [12].
We don't mean to be too flippant about this issue. It is a serious problem
for our hypothesis, and one for which we lack a complete answer. On the
other hand, it isn't all that compelling an argument for a
Calcarea-Homoscleromorpha connection, either.
|
Update: Cross-striated ciliary rootlets have now been reported in the demosponge Asbestopluma, making the rest of this argument somewhat needless. Riesgo, Taylor et al. (2007). |
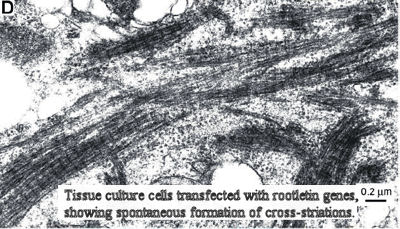
We lack the data draw phylogenetic conclusions based on ciliary rootlets. The problem results, in part, from lack of data. We aren't certain yet what causes the cross-striations. Ciliary rootlets are cable-like systems which attach at one end to the basal body of a cilium (= eukaryotic flagellum) and generally run to a point in the general vicinity of the nucleus, but sometimes elsewhere. The function of rootlets is uncertain. Recently, a group at Harvard has reported that the ciliary rootlets of some vertebrate tissues are probably a homopolymer of a novel protein they have called "rootletin." Yang et al. (2002); Yang et al. (2005). That's Harvard for you. Marvelous work, but they should never be allowed to name things. In any case, rootletin is a more or less distant cousin of myosin, and the cross-striations may result from the alignment of repeating rootletin units. Yang et al. (2002). Ciliary basal bodies are homologues of centrioles and, sure enough, it seems that the centrioles within the nuclei of non-ciliated vertebrate cells may also be joined by strands of rootletin. Yang et al. (2006).
As yet we know nothing about the presence or absence of rootletin in Demospongiae, or for that matter in any sponge. Perhaps rootletin has nothing to do with rootlet cross-striations in sponges. Perhaps it does, but the demosponges (for whatever reason) don't line up the rootletin fibers in a way which exhibits cross-striations. There are not all that many studies of the ciliary rootlets of sponges. Perhaps one or more groups of demosponges have banded ciliary rootlets, at least (like Calcarea), in the embryo and we simply haven't seen them. Finally, sponges react oddly to conventional preparations for electron microscopy. Thus, it is even possible (if unlikely) that the lack of cross-striation in demosponge rootlets is an artifact of preparation.
 Cross-striated rootlets are homoplastic in Eukarya. The real problem with relying on rootlet cross-striation as a synapomorphy is
that the trait is plainly homoplastic in the Eukarya as a whole. For
example, cross-striated ciliary rootlets are also known from
Alveolata, the
heterolobosean amoebae, the parabasalids, the spironemid flagellates, and
Cryothecomonas (a flagellate of uncertain affinities).
Patterson (1999).
This character may also be found in some choanoflagellates, although here the
evidence is less convincing.
Maldonado (2004);
but see
Sperling et al. (2006). In any event, if rootletin turns out to
be the main link between centrioles, it will probably also turn out to be a key
element in organizing mitosis. If so, it is hard to imagine that rootletin
is entirely absent in demosponges. It must simply take on a form which
does not show electron-dense bands in rootlets. Finally, so far as we can
tell, no one claims that any animal completely lacks unstriated
rootlets (i.e. has all rootlets striated). Thus the presence of unstriated
rootlets in Demospongiae is not a synapomorphy, either.
Cross-striated rootlets are homoplastic in Eukarya. The real problem with relying on rootlet cross-striation as a synapomorphy is
that the trait is plainly homoplastic in the Eukarya as a whole. For
example, cross-striated ciliary rootlets are also known from
Alveolata, the
heterolobosean amoebae, the parabasalids, the spironemid flagellates, and
Cryothecomonas (a flagellate of uncertain affinities).
Patterson (1999).
This character may also be found in some choanoflagellates, although here the
evidence is less convincing.
Maldonado (2004);
but see
Sperling et al. (2006). In any event, if rootletin turns out to
be the main link between centrioles, it will probably also turn out to be a key
element in organizing mitosis. If so, it is hard to imagine that rootletin
is entirely absent in demosponges. It must simply take on a form which
does not show electron-dense bands in rootlets. Finally, so far as we can
tell, no one claims that any animal completely lacks unstriated
rootlets (i.e. has all rootlets striated). Thus the presence of unstriated
rootlets in Demospongiae is not a synapomorphy, either.
| Page Back | Page Top | Unit Home | Page Next |