| Palaeos: | Porifera | |
| Porifera | Hexactinellida |
| Page Back | Unit Back | Metazoa | Metazoa Dendrogram | Metazoa References | Pieces | Taxon Index |
| Page Next | Unit Next | Unit Home | Unit Dendrogram | Unit References | Glossary | Time |
PORIFERA |--Archaeocyatha `--+--Stromatoporoidea `--+--Calcarea `--+--Hexactinellida `--+--Demospongiae | |--Tetractinomorpha | `--Ceractinomorpha `--+--Homoscleromorpha `--EUMETAZOA |
 Anatomy Anatomy
Evolution Tales of Silt and Carbon Classification References
|
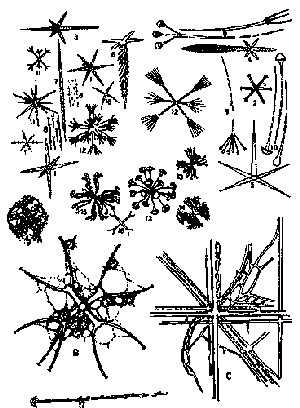 Hexactinellids
and their Spicules: Hexactinellids, commonly called "glass sponges", construct a skeleton composed of simple to complex 6-rayed
siliceous
spicules (left) with a fundamentally
orthogonal
symmetry. In extant heaxactinellids, the spicules are often fused into networks, with the entire structure
appearing as an open mesh or an intricate, basket-like structure. Hexactinellids are exclusively marine, and in the modern oceans are usually found
at depth (generally 450 to 900 m, but up to 5000 m).
Hexactinellids
and their Spicules: Hexactinellids, commonly called "glass sponges", construct a skeleton composed of simple to complex 6-rayed
siliceous
spicules (left) with a fundamentally
orthogonal
symmetry. In extant heaxactinellids, the spicules are often fused into networks, with the entire structure
appearing as an open mesh or an intricate, basket-like structure. Hexactinellids are exclusively marine, and in the modern oceans are usually found
at depth (generally 450 to 900 m, but up to 5000 m).
Some of the anchoring spicules produced by hexactinellids are extraordinarily large. The anchoring spicules of deep-sea sponge Monorhaphis can reach lengths of up to 3 m with a maximum diameter of only 8.5 mm. Hexactinellid spicules are produced in concentric layers -- up to 500 layers in the case of Monorhaphis. Each layer is thus 10-20 μ thick, composed of small grains of silicate within a matrix of undetermined composition (possibly collagen). The silicate layers are built on a core of amorphous silicate. The core, in turn, is built around a square axial channel containing a protein filament about 2 μ in diameter. Müller et al. (2007). Unlike other sponges, hexactinellids form spicules exclusively inside their cells.
Syncytial Organization. So, how can a sponge make giant spicules inside cells? The hexactinellid sponges are said to display the syconoid pattern of organization. To put it mildly, this understates the weirdness of the hexactinellid body plan. In fact, hexactinellids have perhaps the strangest of all animal bodies. Most of the tissue consists of a single giant, multinucleate syncytium. This syncytium forms both "the inner and outer layers of the sponge and is joined by cytoplasmic bridges to uninucleate cellular regions." Leys et al. (2006). That is, a hexactinellid can make giant spicules because its body is composed largely of one giant cell. Compare the Xenophyophorea, gigantic rhizarian protists which have a somewhat similar structure.
This syncytial organization appears to be secondary, as the embryonic
hexactinellid Oopsacas is cellular until well along into gastrula stage
(yes, sponges have a sort of gastrula).
However, the later merger of cells is fairly complete. Even the
micromeres
left at the outer surface are joined to the syncytium by cytoplasmic bridges or
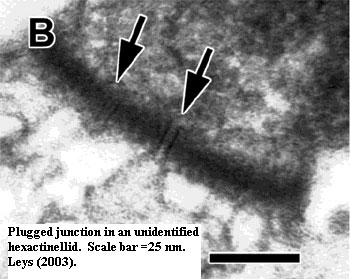
peculiar "plugged junctions" reminiscent of the pores plugged by
Woronin Bodies in the septae between sections of fungal hyphae.
Instead of
choanocytes, hexactinellids have "collar bodies," with collars and
flagellae, but lacking the usual cell bodies. The
 hexactinellids have no
real pinacoderm. The syncytium forms a network of strands -- like a
three-dimensional spider-web, which suspends numerous small chambers, lined with
collar bodies, like the carcasses of so many bugs caught in the web. Each
of these chambers is independently open to the water. The spicule-forming
sclerocytes of the embryo develop in the usual way, but then become
multinucleate and connect to the syncytium by cytoplasmic bridges. Leys et al.
(2006). However, at least in some hexactinellids (Dactylocalyx), some of
these specialized cells apparently remain entirely independent.
hexactinellids have no
real pinacoderm. The syncytium forms a network of strands -- like a
three-dimensional spider-web, which suspends numerous small chambers, lined with
collar bodies, like the carcasses of so many bugs caught in the web. Each
of these chambers is independently open to the water. The spicule-forming
sclerocytes of the embryo develop in the usual way, but then become
multinucleate and connect to the syncytium by cytoplasmic bridges. Leys et al.
(2006). However, at least in some hexactinellids (Dactylocalyx), some of
these specialized cells apparently remain entirely independent.
Anatomy and Physiology -- Leys (2003). Fortunately for us, hexactinellid anatomy and physiology has recently been summarized in an accessible, short review by Leys (2003), for which we provide the following direct link. As usual when we find a really good source of this kind, we will provide only bullet points:
 Early
Hexactinellids had a one-Layer "Quadruled" Skeleton. Hexactinellids have a
reasonably good fossil record, going all the way back before the Cambrian. As
discussed elsewhere, they probably
evolved from the basal radiation of "heteractinids." The fashion for all early sponges was to
maintain a single layer of outer tissue, but increase spicule size with body size. However, it was considered rather
bad form to simply stretch out the old spicule to a new size, as if it were a
Spandex waistband. Rather, the prosperous sponge of Cambrian time grew spicules in an
increasing number of size classes as it expanded, with each size class about 1.3
times larger than the last. Botting & Butterfield (2005).
Early
Hexactinellids had a one-Layer "Quadruled" Skeleton. Hexactinellids have a
reasonably good fossil record, going all the way back before the Cambrian. As
discussed elsewhere, they probably
evolved from the basal radiation of "heteractinids." The fashion for all early sponges was to
maintain a single layer of outer tissue, but increase spicule size with body size. However, it was considered rather
bad form to simply stretch out the old spicule to a new size, as if it were a
Spandex waistband. Rather, the prosperous sponge of Cambrian time grew spicules in an
increasing number of size classes as it expanded, with each size class about 1.3
times larger than the last. Botting & Butterfield (2005).
As it turns out, 1.3 is a convenient number, because (1.3)3 ≈ 2.2. Using every third size class thus lends itself to a structural pattern which Botting & Butterfield call "quadruling." This is illustrated by an actual hexactinellid sponge, Protospongia. Protospongia perfected the quadruling style in the Early Devonian, after which it quite sensibly became extinct. Later hexactinellids grew additional layers of spicule-supported tissue, but abandoned this unique, fractal mode of construction. Id.
Evolutionary History. The earliest sponges identified as hexactinellids are from Ediacaran exposures in Mongolia and India. Tiwari et al. (2000). Hexactinellids were relatively rare in Tommotian Chengjiang (Hagadorn, 2002), but were an important component of the Middle Cambrian Burgess Shale community. Unfortunately, it is not clear whether these earliest discoveries are, in fact Hexactinellida, or part of the basal "heteractinid" radiation. Botting & Butterfield (2005).
 Certainly hexactinellids were important, and perhaps the dominant form of
sponge, from the Middle Cambrian on, into the late Paleozoic (Tiwari et al.,
2000), and occupied much shallower water than modern forms. After the
extinction of protospongeoid, "quadruled" sponges in the Early Devonian, the
hexactinellids evolved in two directions. By the
Late Devonian, hexactine
sponges had evolved a looser skeleton, while hexactinosans (hexactinellid "lithistid" sponges)
created the massively fused siliceous skeleton more typical of Mesozoic and
Cenozoic hexactinellids.
Vishnevskaya
et al. (2002). However, they declined in the
Permian, along with other
sessile
epifauna
and became increasingly restricted to deep waters. Wood (1998). Like
most long-term trends, this one has not been entirely consistent.
Hexactinellids made a considerable, perhaps dominant, contribution to the
Late Jurassic reef system
along the northern borders of the Tethys, extending from Central Asia to the
southeastern United States -- more or less from Tblisi, Georgia to Savannah,
Georgia. Krautter
(1998).
Certainly hexactinellids were important, and perhaps the dominant form of
sponge, from the Middle Cambrian on, into the late Paleozoic (Tiwari et al.,
2000), and occupied much shallower water than modern forms. After the
extinction of protospongeoid, "quadruled" sponges in the Early Devonian, the
hexactinellids evolved in two directions. By the
Late Devonian, hexactine
sponges had evolved a looser skeleton, while hexactinosans (hexactinellid "lithistid" sponges)
created the massively fused siliceous skeleton more typical of Mesozoic and
Cenozoic hexactinellids.
Vishnevskaya
et al. (2002). However, they declined in the
Permian, along with other
sessile
epifauna
and became increasingly restricted to deep waters. Wood (1998). Like
most long-term trends, this one has not been entirely consistent.
Hexactinellids made a considerable, perhaps dominant, contribution to the
Late Jurassic reef system
along the northern borders of the Tethys, extending from Central Asia to the
southeastern United States -- more or less from Tblisi, Georgia to Savannah,
Georgia. Krautter
(1998).
Phylogenetic Position is Uncertain. As discussed at various points elsewhere, the position of the hexactinellids is unclear. The traditional view (to which we still cling) is that hexactinellids are so peculiar that they must have branched very early. Quite possibly, we are making the error of confusing symplesiomorphies with synapomorphies. Yet molecular phylogenies have found nearly every possible grouping of the three major sponge groups and Eumetazoa. See, e.g., Cavalier-Smith & Chao (2003); Rokas et al. (2003); Medina et al. (2001). Hexactinellids, as you might expect, are almost as notorious as nematodes for long-branch artifacts, so the credibility of molecular phylogenies is particularly suspect. Id. The best of this unavoidably bad lot is probably still Schütze et al. (1998), which finds the Hexactinellida to be the earliest-branching sponge group.
 Ecology?. Demosponges have evolved to handle soft substrates and mobile
predators with a variety of subsurface anchors and unpleasant toxins.
Hexactinellids lack these tools. They rely more on tightly woven spicules
and the fact that their soft tissues are rather insubstantial.
Hexactinellids are mostly silicate mesh and empty space. This design also
provides a very high surface to volume ratio. Consequently, it has often
been said that they are able to subsist on dissolved organic carbon, even when
bacteria (the usual sponge fodder) are in scarce supply. On the other
hand, this general design also makes hexactinellids rather sensitive to
sedimentation, which can quickly clog their silicate meshes, or cause them to
become top-heavy and topple over. For similar reasons, hexactinellids need
a relatively low-energy environment. All of these factors favor a deep-water
home, and probably explain why they still do well in such environments.
Krautter (1998). The supposed ability of
hexactinellids to live off dissolved organic carbon
has also led to some elaborate theorizing about the evolution of metazoans. See,
e.g., Sperling
et al. (2006).
Ecology?. Demosponges have evolved to handle soft substrates and mobile
predators with a variety of subsurface anchors and unpleasant toxins.
Hexactinellids lack these tools. They rely more on tightly woven spicules
and the fact that their soft tissues are rather insubstantial.
Hexactinellids are mostly silicate mesh and empty space. This design also
provides a very high surface to volume ratio. Consequently, it has often
been said that they are able to subsist on dissolved organic carbon, even when
bacteria (the usual sponge fodder) are in scarce supply. On the other
hand, this general design also makes hexactinellids rather sensitive to
sedimentation, which can quickly clog their silicate meshes, or cause them to
become top-heavy and topple over. For similar reasons, hexactinellids need
a relatively low-energy environment. All of these factors favor a deep-water
home, and probably explain why they still do well in such environments.
Krautter (1998). The supposed ability of
hexactinellids to live off dissolved organic carbon
has also led to some elaborate theorizing about the evolution of metazoans. See,
e.g., Sperling
et al. (2006).
.. or maybe not ... It therefore came as something of a shock when a very recent empirical study found that most of the last paragraph was myth. Yahel et al. (2007). Hexactinellids are, it seems, even more voracious than most sponges when it comes to removing bacteria from intake water, but they show no signs of interest in organic smoothies. Also, contrary to expectations, they are also partial to fine sediment, which they absorb, lick clean of adhering organics, and spit back out again. To be fair, this is only a single study. But it does illustrate some of the pitfalls of relying on scientific folklore which has never been empirically tested. Sponges are relatively little studied, and a remarkable number of accepted sponge facts turn out to be speculations by some Nineteenth Century worthy, who was simply giving us his best guess, with even less information than we have today.
 We have not attempted to catch up with any recent changes in hexactinellid
taxonomy. There are two main groups (clades or subclasses) and six orders, all of which have a fossil record. The oldest known species, and also the oldest known fossil sponge, is Paleophragmodictya from the Ediacaran of South Australia.
That, and others in the Reticulosa, may or may not be "heteractinids," an
informal taxon of basal sponges with a mosaic of demosponge, calcarean, and
hexactinellid characters.
We have not attempted to catch up with any recent changes in hexactinellid
taxonomy. There are two main groups (clades or subclasses) and six orders, all of which have a fossil record. The oldest known species, and also the oldest known fossil sponge, is Paleophragmodictya from the Ediacaran of South Australia.
That, and others in the Reticulosa, may or may not be "heteractinids," an
informal taxon of basal sponges with a mosaic of demosponge, calcarean, and
hexactinellid characters.
Hexactinellida
Amphidiscophora
Reticulosa (extinct) (Ediacaran - Permian)
Hemidiscosa (extinct) (Carboniferous - Cretaceous)
Amphidiscosa / Amphidiscosida (Ordovician - Present)
Hexasterophora
Lyssacinosa / Lyssacinosida (Ordovician - Present)
Hexactinosa / Hexactinosida (Devonian - Present)
Lychniscosida (Triassic - Present)
Classification according to
Fossil Record of the Hexactinellida, Valid PMPD Orders and
Synopsis of the described taxa of the world
Since the Reticulosa are presumably paraphyletic, the Amphidiscophora are also questionable. Accordingly, for working purposes, we draw the cladogram like this:
Porifera (Heteractinida)
|--Eifellia
`--+--Hexactinellida
| |--Protospongoidea
| `--+--Hemidiscosa
| |--Amphidiscosa
| `--Hexasterophera
`--+--Demospongiae
`--+--Calcarea
`--EUMETAZOA
 |
 |
 |
Brachiospongia digitata (Owen)
Trenton Limestone = ?Sandbian (Ordovician) Kentucky, USA. (Laurentia) diameter about 8 or 10 cm order Lyssakida - family Brachiospongiidae image © C. L. & M. A. Fenton, The Fossil Book, p.64
|
Hydnoceras tuberosum Conrad
Chenmung = Late Frasnian (Devonian) height about 12 cm New York (Western/Central Euramerica) image © xxxx
|
Titusvillea drakei Caster
A colonial glass sponge Early Mississippian = Tournaisian height of this specimen about 12 cm Cussewago Beds, northwestern Pennsylvania (Western/Central Euramerica) image © C. L. & M. A. Fenton, The Fossil Book, p.64
|
| Page Back | Page Top | Unit Home | Page Next |