| Palaeos: | Demospongiae | |
| Porifera | Demospongiae - 1 |
| Page Back | Unit Back | Metazoa | Metazoa Dendrogram | Metazoa References | Pieces | Taxon Index |
| Page Next | Unit Next | Unit Home | Unit Dendrogram | Unit References | Glossary | Time |
PORIFERA |--Archaeocyatha `--+--Stromatoporoidea `--+--Calcarea `--+--Hexactinellida `--+--Demospongiae | |--Tetractinomorpha | `--Ceractinomorpha `--+--Homoscleromorpha `--EUMETAZOA |
Introduction Spicules: a New Look Sponge Skin Rootlets and Relationships New Developments History The Sequence of Sponges
|
 It is not a good sign when sponge experts cannot
agree on the name of the most common large taxon of sponges.
"Demospongiae" is by far the majority choice. However, there are some who
insist on "Demospongia," (e.g.,
Medina et al., 2001;
and Benton & Harper, 1997
-- sometimes) or even "Demospongea" (Benton & Harper, 1997 -- the rest of the
time). This may be one of those Linnaean things, with deep significance for those
concerned with taxonomic rank. It turns out that all of these spellings are good Latin.
The -ea and -ia forms are alternate spellings of the singular,
which is the actual (if unlikely) Latin word for sponge: spongia, or
spongea, as the case may be. Still, the ambiguity portends problems;
and there are a great many of them.
It is not a good sign when sponge experts cannot
agree on the name of the most common large taxon of sponges.
"Demospongiae" is by far the majority choice. However, there are some who
insist on "Demospongia," (e.g.,
Medina et al., 2001;
and Benton & Harper, 1997
-- sometimes) or even "Demospongea" (Benton & Harper, 1997 -- the rest of the
time). This may be one of those Linnaean things, with deep significance for those
concerned with taxonomic rank. It turns out that all of these spellings are good Latin.
The -ea and -ia forms are alternate spellings of the singular,
which is the actual (if unlikely) Latin word for sponge: spongia, or
spongea, as the case may be. Still, the ambiguity portends problems;
and there are a great many of them.
Here, we concentrate on a few areas which have generated a pile of recent papers. We thought they might be useful for phylogenetic purposes. As you may have gathered, sponge phylogeny is currently in a sorry state. In fact, we even found ourselves nodding and smiling at Maldonado (2004), whose ideas approach the downright weird. Maldonado argues that choanoflagellates may be descended from demosponges, rather than vice-versa. Weird or not, Maldonado makes a plausible case for the reverse arrangement. In fact, this scheme solves certain otherwise intractable problems in sponge phylogeny. For example, the fossil record (and other evidence, discussed later) suggests that carbonate skeletons came first, followed by silicate spicules. Botting & Butterfield (2005). However, as Maldonado points out, some choanoflagellates produce silicate loricas (Cavalier-Smith & Chao, 2003) [11]. Perhaps they inherited this ability from demosponge ancestors.
Not that we agree with Maldonado. It is not entirely clear that even Maldonado really agrees with Maldonado. Rather, he seems to be telling us, all too skillfully, to avoid the facile or "consensus" answers. Dissonance and contradiction lie in ambush for the hopeful sponge taxonomist. We could not evade all of these traps, but perhaps our mistakes may serve as a warning to others.
We will deal here with a subset of the issues -- spicule formation, development, and some associated biochemistry. These seem to lead to a phylogeny which mostly agrees with the fossil record, as interpreted by Botting & Butterfield (2005), but places the Eumetazoa on the demosponge branch. The track is tortuous and uncertain, with frequent excursions into dead-ends and suppositions, pasted together with guesswork. While this is not all that different from the rest of Palaeos, perhaps, it is even messier.
![]() Classically, Demospongiae was
a taxon held together by
spicules. Almost anything sponge-like, with
siliceous spicules, was deemed to be a
demosponge. Needless to say,
life has become considerably more complicated. To form their skeletal
elements, different demosponges use
varying combination of
spongin,
silica, calcite,
or even
aragonite -- or even none of the above, since
a few demosponges have no skeleton or, like Americans, bulk up on debris without much regard for
composition. The collagen protein spongin may prove to be a true
synapomorphy
of the Demospongiae. But even this is uncertain, because any sponge with spongin is
assumed to be a demosponge. Most workers with a sophisticated knowledge of
sponges -- and who would otherwise know better -- likewise assume that the molecule folks will take care of phylogeny,
and have stopped writing about the phylogenetic implications of their results. Unfortunately, as we will see, sequence-based
molecular phylogenies have little credibility for high-level sponge taxa.
Classically, Demospongiae was
a taxon held together by
spicules. Almost anything sponge-like, with
siliceous spicules, was deemed to be a
demosponge. Needless to say,
life has become considerably more complicated. To form their skeletal
elements, different demosponges use
varying combination of
spongin,
silica, calcite,
or even
aragonite -- or even none of the above, since
a few demosponges have no skeleton or, like Americans, bulk up on debris without much regard for
composition. The collagen protein spongin may prove to be a true
synapomorphy
of the Demospongiae. But even this is uncertain, because any sponge with spongin is
assumed to be a demosponge. Most workers with a sophisticated knowledge of
sponges -- and who would otherwise know better -- likewise assume that the molecule folks will take care of phylogeny,
and have stopped writing about the phylogenetic implications of their results. Unfortunately, as we will see, sequence-based
molecular phylogenies have little credibility for high-level sponge taxa.
The classical sponge taxonomies were based largely on spicule morphology. This was ultimately found to be an unsatisfactory tool for phylogeny. However, spicules have also recently become the subject of some serious molecular biology. These results suggest new approaches to phylogeny. Spicules are undeniably tempting as tools for sponge taxonomy because there are so many of them. Demosponges and hexactinellids may be over 75% spicule by dry weight. Müller et al. (2006); Barthel (1995). (Of course, there are not many dry sponges -- but it's just the kind of specious, pseudo-quantitative drivel you can throw around to amaze your friends.)
Silica Transport. Now, all of this spicule mass has to come from somewhere. The concentration of silica
in sea water is highly variable, but typically less than
50µM.
Abe & Watanabe
(1992). If we have kept all our decimal points in
 order, this means that a
sponge must often suck all the available silicate out of almost 1000
liters of seawater to make a gram of silicate spicule. Carbonate, by
contrast,
is about 5-40 times more plentiful than silicate -- at least in modern oceans. Uriz
(2006).
More to the point, carbonate can be produced from carbon dioxide, a by-product
of the metabolic activity of the sponge and its bacterial symbionts. This
is yet
another pseudo-quantitative factoid, but it raises the following
interesting question: if it takes considerably more work to collect silicates than carbonates, would this support the idea (Botting
& Butterfield, 2005) that carbonate skeletons probably evolved first?
order, this means that a
sponge must often suck all the available silicate out of almost 1000
liters of seawater to make a gram of silicate spicule. Carbonate, by
contrast,
is about 5-40 times more plentiful than silicate -- at least in modern oceans. Uriz
(2006).
More to the point, carbonate can be produced from carbon dioxide, a by-product
of the metabolic activity of the sponge and its bacterial symbionts. This
is yet
another pseudo-quantitative factoid, but it raises the following
interesting question: if it takes considerably more work to collect silicates than carbonates, would this support the idea (Botting
& Butterfield, 2005) that carbonate skeletons probably evolved first?
We thought we could shore up this speculation with a quick peek at transporter mechanisms, but we were wrong. As the figure from Schröder et al. (2004) shows, carbonate and silicate are imported from seawater using the same transporter. The assiduous reader may recall that this is not the only commonality between these spicule systems. Silicase, the enzyme that dissolves silicate and helps shape the spicules, is essentially the same as carbonic anhydrase, the enzyme which creates carbonate from carbon dioxide. Müller et al. (2007). Interestingly, eumetazoan homologues of this same transporter family are frequently associated with carbonic anhydrase, combining the two functions to regulate pH. Pushkin & Kurtz (2006). Whether this association between carbonate-silicate transporter and carbonic anhydrase also occurs in sponges is not known.
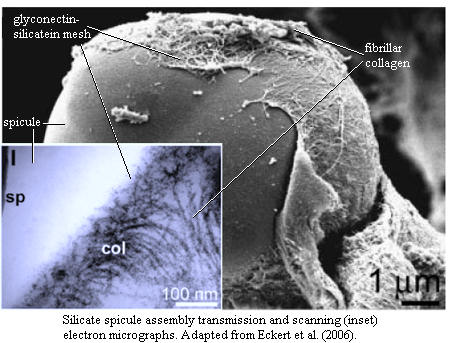
Spicule Assembly. Demosponges actually assemble spicules using a rather complex process
described by Schröder
et al. (2004) and
Eckert et al.
(2006). Parenthetically, Schröder
et al. (2004) makes it onto our very short list of
Amazing Sponge Papers,
which we recommend reading at this
direct link.
Very briefly, the spicule is nucleated inside a cell vacuole on a core of
silicatein
overlain with a meshwork of
glyconectin.
After the spicule is a few µ long, it is extruded into the
mesohyl.
There it is surrounded, but never contacted, by fibrillar
collagen. The
collagen apparently serves to pattern additional net-like of meshes of
glyconectin and, presumably, additional
 silicatein, which adds additional
layers.
silicatein, which adds additional
layers.
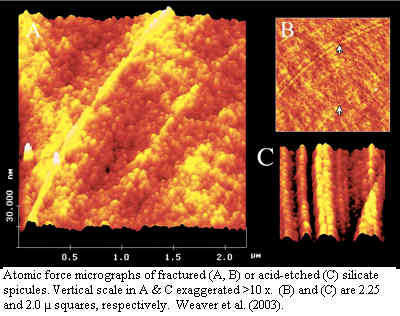
Remarkably, this work is consistent with even more detailed structural predictions made using low-angle X-ray scattering (Croce et al., 2004) , atomic force microscopy (Weaver et al., 2003) and magnetic resonance imaging (MRI) (Müller et al., 2006). For our purposes, the most important fact that all this stuff tells us is this: the component silicates are not at all crystalline. They are laid down as blobs of amorphous silicate trapped on grids of glycoproteins, the glyconectins. The blobs seems to be arranged carefully, in concentric layers, exactly one blob thick, and in a manner which does not permit adjacent blobs to merge.
Comparison with other Sponge Groups. How does demosponge spicule development compare with spicule formation in other sponge classes? It's a bit hard to tell, since no one has looked at the other groups in this kind of detail. The system in Calcarea is poorly known, but also seems to involve collagen fibrils densely packed around the growing spicule. However, unlike siliceous spicules, calcareous spicules are essentially crystalline and include comparatively little protein of any kind. The growing calcite spicule does have some sort of proteinaceous sheath which closely controls crystal growth, but the sheath is displaced by the growing spicule -- not incorporated. Weiner & Addadi (1997), Botting & Butterfield (2005). See also Uriz (2006).
Hexactinellids may produce spicules in essentially the same fashion as demosponges. However, hexactinellid spicules include a higher level (~15%) of organic materials than demosponge spicules (~10%), and the silicate blobs are less tightly packed. Croce et al., 2004. Maldonado & Riesgo (2007) have recently examined the formation of unusual "intra-epithelial" spicules in a homoscleromorph sponge. These spicules are not typical, being located inside certain epithelial cells. However, we do not see their description of spicule synthesis as being fundamentally different from the demosponge system, except that inner protein layers are more evident.
 The
Primitive Condition. With
this background, we may repeat Botting & Butterfield's important question. Which
is primitive for sponges, silicate spicules or carbonate spicules? Like
most important choices in paleo or politics, we suspect that the dichotomy is false and the
correct answer is "none of the above." Spicules are not primitive for sponges.
Massive carbonate skeletons are primitive for sponges, as found in the
Archaeocyatha and stromatoporoids. In any case, massive calcareous
skeletons are primitive for nearly every other group which has evolved a
mineralized skeleton. Why not sponges?
The
Primitive Condition. With
this background, we may repeat Botting & Butterfield's important question. Which
is primitive for sponges, silicate spicules or carbonate spicules? Like
most important choices in paleo or politics, we suspect that the dichotomy is false and the
correct answer is "none of the above." Spicules are not primitive for sponges.
Massive carbonate skeletons are primitive for sponges, as found in the
Archaeocyatha and stromatoporoids. In any case, massive calcareous
skeletons are primitive for nearly every other group which has evolved a
mineralized skeleton. Why not sponges?
Spicules vs. Skeletons. But massive carbonate skeletons are not carbonate spicules. Chemically, they could scarcely be more different and still be composed of the same ground substance. The carbonate spicules of Calcarea are essentially single crystals whose orientation seems to be controlled on a nanometer scale. Calcite crystallizes spontaneously under certain, biologically relevant conditions. A calcarean sponge needs no "carbonatein" enzyme, by analogy the silicatein of demosponges, to encourage the self-assembly of spicules; but the direction of growth must be carefully controlled.
By contrast, massive carbonate skeletons are composed of amorphous calcite. This requires a mechanism to avoid crystallization. Sponges do this by aggregating granules of amorphous calcite. This calcite is kept amorphous by an admixture of glycoproteins. Otherwise, it would transform into a crystalline morph very quickly. Weiner & Addadi (1997).
Phylogenetic Consequences. If we have this correct, the probable sequence of events is not difficult to reconstruct. Stem group sponges (archaeocyaths, Paleozoic stromatoporoids) had amorphous calcite skeletons. Subsequently one lineage (at least) developed spicules, radiating into a number of different types (e.g. Eifellia, chancelloriids). Note that amorphous calcite can be used to make spicules, as in the extant urochordate Pyura.
These early spiculate sponge groups probably used a variety of different "recipes" for making spicules. Some recipes included silicate. The use of silicate actually required little change because, as discussed, the silica and carbonate mechanisms for spicule construction have much in common. Ultimately, two spiculate groups dominated. One, the Calcarea, continued to use carbonate, but adapted to manipulate crystal growth, gradually reducing the expensive investment in glycoprotein additives. The other branch, including demosponges and hexactinellids, kept the basics of the amorphous mineral plus glycoprotein system, but switched to the structurally stronger and more stable silicates. Both groups retained the basic equipment for making massive calcareous skeletons, a trait which pops up now and again in both lineages.
Plausible? Let's see how it fits with other facts, and, if possible, whether it allows us to place the Eumetazoa somewhere...
| Page Back | Page Top | Unit Home | Page Next |