Chlorobionta (Green Plants) - 2
We will cover the higher taxa of lower plants in two blocks: Chlorobionta and
Embryophyta. The prasinophytes (basalmost chlorobionts), chlorophytes and
charophytes are essentially algae, which normally impinge on our consciousness
just long enough to apply a little wasabi and shoyu. Arigato,
and next I'll have ni unagi, kudasai. Don't try that with an
embryophyte. There's a differnce between sushi and soba.
Embryophytes are mostly land plants, and it was the ability of plants to live on
land that allowed all the other branches of life to live on land as well.
In fact, only the plants can really be said to have adapted to land. With
few exceptions, the rest of life simply adapted to plants.
 The general characteristics of the green plants
are touched on above. The purpose of this section is to
introduce the prasinophytes. These are a paraphyletic group of green algae which
radiate from the base
of the Chlorobionta. Most are photosynthetic
flagellates. In addition, the prasinophytes are the only mixotropic
plants, i.e., they obtain food both by photosynthesis and
phagotrophy. This is, presumably, how they obtained
chloroplasts in the first place. The general characteristics of the green plants
are touched on above. The purpose of this section is to
introduce the prasinophytes. These are a paraphyletic group of green algae which
radiate from the base
of the Chlorobionta. Most are photosynthetic
flagellates. In addition, the prasinophytes are the only mixotropic
plants, i.e., they obtain food both by photosynthesis and
phagotrophy. This is, presumably, how they obtained
chloroplasts in the first place.
The phycomate prasinophytes (those
with large, thick-walled floating stages, or
"phycomata") have received special attention because
of their extremely long fossil record. Phycomata are known as acritarchs well into Proterozoic
time. One genus (Tasmanites) dates back to 600
Mya. Javaux
et al. (2004) have turned up an entire menagerie of
forms from the
Mesoproterozoic,
and even beyond (at least 1500 Mya), which are almost certainly
eukaryotic and could well be prasinophytes, or somewhat stemward
of the plants. They cannot be too distantly related, as
the presence of thick organic walls, with extreme resistance to
degradation, seems to be a trait of the plant-chromist lineage.
One of these in particular, Leiosphaeridia crassa, from
the c. 1460 Mya Roper Fm. of northern Australia, is being
investigated as a possible green alga. Interestingly, in Recent
or merely Paleozoic forms, these relatively large, thick-walled
morphs are associated with moderately anoxic conditions and
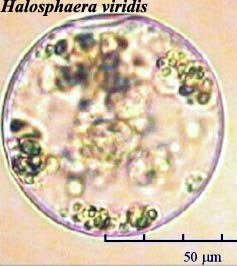
nutrient exhaustion during algal blooms. Image: Halosphaera viridis
from the National
Oceanographic Data Center.
ATW041212. Text public domain. No rights
reserved.
|
|
| Chlorobionts |
 Within the Chlorobionta are two
large clades making up the "green algae." The
green algae, as currently conceived, have no formal taxonomic
name. We will define the group as Quercus + Chlamydomonas.
The corresponding stem clades are Chlorophyta (Chlamydomonas
> Quercus) and Charophyta (Quercus >
Chlamydomonas). "Chlorophyta" is also the old name for
all green algae, so this is perhaps unnecessarily
confusing. Tough luck. The ambiguity is now so
embedded in the literature that there's nothing anyone can do
about it. Within the Chlorobionta are two
large clades making up the "green algae." The
green algae, as currently conceived, have no formal taxonomic
name. We will define the group as Quercus + Chlamydomonas.
The corresponding stem clades are Chlorophyta (Chlamydomonas
> Quercus) and Charophyta (Quercus >
Chlamydomonas). "Chlorophyta" is also the old name for
all green algae, so this is perhaps unnecessarily
confusing. Tough luck. The ambiguity is now so
embedded in the literature that there's nothing anyone can do
about it.
The Chlorophyta have largely been delineated by molecular
techniques, so it is a bit difficult to describe their
characters. We know of two possible synapomorphies of the
Chlorophyta. First, chlorophyte sexual forms bear paired
apical flagellae usually separated by 180°, but sometimes at
the same end. Second, they retain the nuclear envelope
during mitosis. Indeed, chlorophytes seem to be
distinguished by a variety of bizarre variations on the usually
pedestrian theme of mitosis; however those variations are not entirely consistent within the group.
Like the land plant lineage, they tend to form large
aggregates, with some tissue differentiation (primarily
holdfasts and reproductive structures). They are very
often found in terrestrial and fresh water environments, with a
distinct preference for very cold environments, such as under
snow cover, or even within Antarctic ice. Various species
are important in forming symbiotic relationships with fungi, i.e.,
lichens. As with all green algae, chlorophytes tend to
have a double cell wall -- an inner wall of cellulose
and an outer gelatinous wall of protein, particularly pectin,
known in higher plants as a marker for parenchyma.
Starch stored in pyrenoids,
located inside the chloroplasts.
Image: Ulva lobata from the California
Biota Website.
ATW041212. Text public domain. No rights
reserved.
|
|
 The
Charophyta are the other lineage of green algae, the group which
includes the land plants. Karol
et al. (2001). As mentioned above, our working
definition is Quercus (oak) > Chlamydomonas.
The Charophyta have recently been referred to as the Streptophyta,
but the reasons
given for this change in nomenclature are probably
insufficient. Unfortunately, the name is also frequently,
and wrongly, used in place of Charophycea or Charales to
describe the stoneworts -- one of several distinct groups of
charophytes. The
Charophyta are the other lineage of green algae, the group which
includes the land plants. Karol
et al. (2001). As mentioned above, our working
definition is Quercus (oak) > Chlamydomonas.
The Charophyta have recently been referred to as the Streptophyta,
but the reasons
given for this change in nomenclature are probably
insufficient. Unfortunately, the name is also frequently,
and wrongly, used in place of Charophycea or Charales to
describe the stoneworts -- one of several distinct groups of
charophytes.
The synapomorphies
of the group are said to include the the dissolution of the
nuclear membrane during mitosis and the presence of paired
flagella (when flagella are present at all) directed
perpendicularly to each other. In addition, the
charophytes are strongly inclined toward growth as long
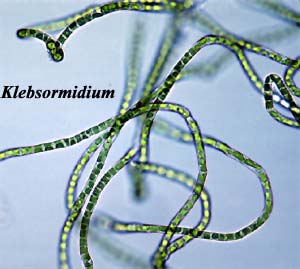
filaments. Image: Klebsormidium
from The
Delwiche Lab at the Univ. of Maryland. ATW041212. Text public domain. No rights
reserved. |
|
| Charophytes |
 The Embryophyta constitute the terrestrial or land plants, the first representatives of which appeared during
the Silurian or possibly even the
Middle or Late Ordovician period. The most primitive of these are nonvascular land plants, a group that
classically includes liverworts (Hepatophyta / Hepaticopsida), hornworts (Anthocerotophyta / Antheroceratopsida) and
mosses
(Bryophyta). The majority of land plants however are included within the huge and diverse clade awkwardly named
Tracheophyta, including the Vascular Pants, and a basal radiation of plants loosely called rhyniophytes. The Embryophyta constitute the terrestrial or land plants, the first representatives of which appeared during
the Silurian or possibly even the
Middle or Late Ordovician period. The most primitive of these are nonvascular land plants, a group that
classically includes liverworts (Hepatophyta / Hepaticopsida), hornworts (Anthocerotophyta / Antheroceratopsida) and
mosses
(Bryophyta). The majority of land plants however are included within the huge and diverse clade awkwardly named
Tracheophyta, including the Vascular Pants, and a basal radiation of plants loosely called rhyniophytes.
We treat Embryophyta in a specialized sense,
as Quercus + moss. This may be a mistake, as this
definition probably excludes the liverworts (see image) and
perhaps even the hornworts. Both of these groups have
traditionally been thought of as embryophytes.
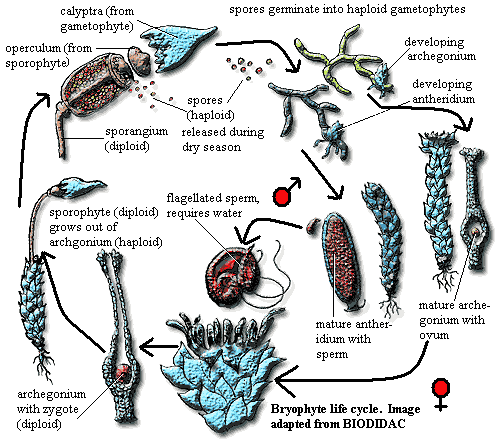
Embryophytes (including liverworts) have the following synapomorphies:
1) a life cycle with alternation
of generations 2) apical cell growth (some kind of meristem-like
growth organization), 3) cuticle
(needed to control water loss on land), 4) antheridia (male gametophyte organs),
and 5) archegonia (female gametophyte
organs). The more derived embryophytes are vascular
plants. Vascular plants have an elaborate system of conducting cells, consisting of xylem - in which water and minerals are transported) and
phloem (in which carbohydrates are transported). This method of internal
support enables them to stand and grow upright and pull up nutrients against
the force of gravity. There are two developmental grades - those
that reproduce by means of spores, and hence are dependent on water or
extensive moisture (e.g. ferns), and those that reproduce by means of seeds
(e.g. conifers and flowering plants). The most primitive forms reproduce by means of spores
(haploid
(1N) spores). They generally require a moist environment, because the
flagellated sperm require water for fertilization.
text by MA Kazlev 2002, revised ATW041215, ATW090227.
|
|
The Embryophytes, then, are plants with an alternation of
generations and some ability to live on land. The basal embryophytes were
still not land plants, since they required, and still require, open water to
propogate. As we define the Embryophyta, they split basally into mosses (Bryophyta)
and land plants (Tracheophyta). The tracheophytes include a basal miscellany of extinct "rhyniophytes"
and two important living groups: the Lycophytina (lycopsids and the extinct zosterophylls)
and the Euphyllophyta (ferns and seed plants).
Embryophyta
|
|
Embryophyta
|
 If
the mosses had not survived into the present, we would be forced
to invent them as just the sort of intermediate we might expect
between essentially aquatic algae and fully terrestrial
plants. Mosses do have differentiated stems.
Although these are generally only a few millimeters tall, they
are still designed to provide mechanical support against gravity
without help from water -- the first such structure in any
kingdom. Bryophytes
also have leaves. These are typically one cell thick and
lack veins, although they may have a central thickening for
support. Mosses also have rhizomes.
These may have some function in extracting soil nutrients,
although their primary function seems to be mechanical
attachment to the substrate. Thus they are not true roots,
but do approach that condition. If
the mosses had not survived into the present, we would be forced
to invent them as just the sort of intermediate we might expect
between essentially aquatic algae and fully terrestrial
plants. Mosses do have differentiated stems.
Although these are generally only a few millimeters tall, they
are still designed to provide mechanical support against gravity
without help from water -- the first such structure in any
kingdom. Bryophytes
also have leaves. These are typically one cell thick and
lack veins, although they may have a central thickening for
support. Mosses also have rhizomes.
These may have some function in extracting soil nutrients,
although their primary function seems to be mechanical
attachment to the substrate. Thus they are not true roots,
but do approach that condition.
The bottom line is that, structurally, mosses
really differ from rhyniophytes in only one aspect: mosses lack
specialized vascular
tissues. That alone is sufficient to explain the lack of
big leaves, long stems, and true roots. This whole complex
of characters is thus probably primitive. The other
distinctive character of mosses is that the plant we normally
observe is the haploid,
gametophyte
stage. But this character is shared with liverworts (basal
embryophytes) and so is also probably plesiomorphic.
Curiously, in hornworts (also basal
embryophytes) the sporophyte generation is dominant. In
addition, it turns out that the leaves of moss probably evolved
independently from the leaves of higher plants. So the
relationships of the mosses and basal embryophytes are still
uncertain. What really does seem to set mosses apart is
their unique form of leaf. What really seems to unite
mosses with higher plants is (a) the presence of stomata
to control water loss and (b) meristem
(apical growth) in the sporophyte generation. See, Friedman
et al. (2004). Phylogenetically, we treat
Bryophyta as Moss > Quercus.
Image: adapted from BIODIDAC.
ATW041215. Public domain. No
rights reserved.
|
|
 See Tracheophyta.
That section covers the basal rhyniophytes, such as Horneophyton,
which were the first real land plants. These probably
evolved in the Ludlow
and formed the stem group for all other land plants.
Consequently, they are paraphyletic. The working phylogenetic definition is definition is Quercus > moss. Cantino et al. (2007). See Tracheophyta.
That section covers the basal rhyniophytes, such as Horneophyton,
which were the first real land plants. These probably
evolved in the Ludlow
and formed the stem group for all other land plants.
Consequently, they are paraphyletic. The working phylogenetic definition is definition is Quercus > moss. Cantino et al. (2007).
This
group is characterized by the ability to reproduce without open
water. Anatomically, in all tracheophytes, the (diploid) sporophyte
generation is dominant, and the sporophyte is branched.
For this reason, these plants are often referred to as
polysporangiophytes. In addition, the archegonium
develops inside the body of the plant, rather than being
superficial as in mosses and most basal embryophytes. Kenrick
& Crane (1997).
Horneophyton
and a few other basal forms lack tracheids.
That is, they are avascular plants. However, almost all
other tracheophytes have some development of specialized vascular
tissues. The most basal tracheid type, present in most stem
tracheophytes, appears to be the S-type
tracheid. Image: Horneophyton
sculpture by Stephen Caine for the Rhynie
Research Group, University of Aberdeen. Image
substantially modified by ATW041229.
ATW041229. Modified ATW090227.
Text public domain. No rights reserved. |
|
| Tracheophyta |
The Lycophyta include
the lycopsids, zosterophylls,
and related forms, including (probably) a number of plants often
treated as basal rhyniophytes, such as Baragwanathia.
Kenrick
& Crane (1997). Since they are a complex group and
are treated extensively elsewhere,
we will defer discussion to a revision of the existing
materials.
|
|
The clade that unites
oak trees and ferns is Euphyllophyta = Quercus + Equisetum. The two complementary
stem clades are Monilophyta and Spermatophytata.
Euphyllophytes are characterized (Kenrick
& Crane, 1997) by monopodial
or pseudomonopodial
branching, helical arrangement of branches, small, pinnule-like
vegetative branches, the branch apex is recurved or coiled,
paired sporangia
which split open along one side through a single slit, and
radially-alligned xylem
in the larger axes. Only early Euphyllophytes have P-type
tracheids. Kenrick & Crane identified this
clade based entirely on morphological characters. However,
Euphytophytina has also been recovered, with essentially the
same structure, using ssu
rDNA. Duff
& Nickrent (1999). ATW041216
Text public domain. No rights reserved. |
|
|
|
|
Euphyllophytes
|
 The Monilophyta are the
horsetails and ferns, including the Psilotidae (whisk ferns). They are closely related to the seed
plants. Pryer
et al. (2001). So, for example, they exhibit
apical growth (meristem)
in both sporophyte
and gametophyte
generations. They have well-developed roots megaphyllous
leaves and the vascular
system needed to make use of both. However, both may have
been evolved independently of higher plants. Friedman
et al. (2004). In addition, Monilophyta
lack a complete vascular
cambium, and growth of xylem is restricted to lobes of
the primary
xylem strand. The Monilophyta are the
horsetails and ferns, including the Psilotidae (whisk ferns). They are closely related to the seed
plants. Pryer
et al. (2001). So, for example, they exhibit
apical growth (meristem)
in both sporophyte
and gametophyte
generations. They have well-developed roots megaphyllous
leaves and the vascular
system needed to make use of both. However, both may have
been evolved independently of higher plants. Friedman
et al. (2004). In addition, Monilophyta
lack a complete vascular
cambium, and growth of xylem is restricted to lobes of
the primary
xylem strand.
Since this
is a new clade -- discovered, for all practical purposes, by
Preyer's group, we have little to say about Monilophyta as a
taxon, and defer discussion to a fuller consideration of its
three component parts. The Psilotidae are the most basal,
followed by the horsetails, then the remainder of the ferns. We
apply a crown group defiition to Monilophyta: Equisetum
+ ferns. Image:
Psilotum nudum from the University
of Wisconsin - Madison Plant Systematics Collection. ATW041216.
Text public domain. No rights reserved.
|
|
|
|
|
 The clade that unites oaks
and lycopsids is Eutracheophyta. The two complementary
stem clades are Lycophyta
and Spermatophytata = Quercus > Lepidodendron.
A second way to look at Spermatophytata is as the stem group
leading to angiosperms. It includes Trimerophyta and the
progymnosperms, in fact everything up to and including the seed
plants (Spermatopsida).
However, we will only be concerned with the more basal forms
for now. A third way of considering Spermatophytata
is as the seed
plants. However, this applies only to living forms.
The basal Trimerophyta and their immediate descendants (assuming
Trimerophyta is paraphyletic)
lacked seeds, true leaves, or even, perhaps, roots. It is
quite likely that virtually all the important land plant
adaptations were independently developed in the monilophyte
and spermatophytate lineages. The clade that unites oaks
and lycopsids is Eutracheophyta. The two complementary
stem clades are Lycophyta
and Spermatophytata = Quercus > Lepidodendron.
A second way to look at Spermatophytata is as the stem group
leading to angiosperms. It includes Trimerophyta and the
progymnosperms, in fact everything up to and including the seed
plants (Spermatopsida).
However, we will only be concerned with the more basal forms
for now. A third way of considering Spermatophytata
is as the seed
plants. However, this applies only to living forms.
The basal Trimerophyta and their immediate descendants (assuming
Trimerophyta is paraphyletic)
lacked seeds, true leaves, or even, perhaps, roots. It is
quite likely that virtually all the important land plant
adaptations were independently developed in the monilophyte
and spermatophytate lineages.
What
seems to have set Spermatophytata apart quite early is not, in
fact, the development of seeds, but the evolution of a full vascular
cambium which permitted secondary
growth. Early plants with apical growth were
able to use that trait to grow taller and (a) get more sunlight
(b) shade their competition and (c) have a better shot at spore
dispersal. However, supporting a long stalk is much easier
with a wider central column. Less derived groups either
had no way to do this, or developed lateral lobes of the apical meristem.
The latter worked, but required the tree to grow wide before
it grew tall. The evolution of a complete vascular cambium
permitted the tree to grow just wide enough to suit its height
-- growing continuously wider as it grew tall. The
evolution of seeds follwed this innovation. Seeds
are embryonic sporophytes,
held in a sort of metabolic stasis and provided with enough food
to get started once their growth has been re-stared by exposure
to suitable growing conditions. Well adapted seeds
combined sexual reproduction with spore-like wide dispersal and
so made the alternation
of generations obsolete. However, early seeds, which
might lack these refinements, probably evolved on tall trees
which gave any sort of propagule
a head start in dispersal. Image:
Psilophyton from the PhD
project page of Ben Sheppard at the University of
Sheffield. ATW041216
Text public domain. No rights reserved. |
|
The Spermatophytata are the stem group for our next major
division, the Spermatopsida.
 Introduction to the Plantae - The green kingdom
Introduction to the Plantae - The green kingdom

 Integrative Biology 181/181L - Paleobotany - at UC Berkeley - excellent!!!!
Integrative Biology 181/181L - Paleobotany - at UC Berkeley - excellent!!!!

 Land Plants On-line - covers recent plants only, links to images etc
Land Plants On-line - covers recent plants only, links to images etc
 International Plant Taphonomy Meeting - The purpose of the International Plant Taphonomy Meetings is to stimulate scientific research and to promote contacts among scientists engaged in the study of plant taphonomy including living and fossil plants of all geological periods.
International Plant Taphonomy Meeting - The purpose of the International Plant Taphonomy Meetings is to stimulate scientific research and to promote contacts among scientists engaged in the study of plant taphonomy including living and fossil plants of all geological periods.
 Botany
Web Sites by Subject - excellent annotated list of links to Botany and related subjects - note, some of these links are no longer current
Botany
Web Sites by Subject - excellent annotated list of links to Botany and related subjects - note, some of these links are no longer current
A BASIC BIOLOGICAL CLASSIFICATION OF PLANT-LIKE ORGANISMS
 A History of Palaeozoic Forests - Hans Kerp - very informative - originally published in German. Deals with forests of the Devonian, Carboniferous, and Permian periods.
A History of Palaeozoic Forests - Hans Kerp - very informative - originally published in German. Deals with forests of the Devonian, Carboniferous, and Permian periods.
Hans' Paleobotany Pages - info on the earliest land plants and on the lycopod Lepidodendron
Carboniferous Forests - Ralph E. Taggart - good non-technical intro, covers main groups of Carboniferous plants, also brief mention of insects, amphibians, and reptiles
The Biota of Early Terrestrial Ecosystems: The Rhynie Chert - includes useful information on Early Devonian plants from this location
The
First Land Plants - The Conquest of the Land - gives a good introduction to basic concepts regarding the transition of plants from water to land
Orto Botanico - somewhat technical but not too difficult coverage of plants and paleobotany. Includes glossary.
Integrative Biology 181/181L - Paleobotany - at UC Berkeley - includes material on Paleozoic plants. A bit technical but if you stick at it you will learn a lot.
images not loading? | error messages? | broken links? | suggestions? | criticism?
Page last modified ATW090227 MAK050103
Checked ATW050503