 Litopterans and Leprechauns
Litopterans and Leprechauns| Meridiungulata | ||
| The Vertebrates | Didolodonts & Litopterns |
| Vertebrates Home | Vertebrate | Vertebrate |
|
Abbreviated Dendrogram
Eutheria
│
└─Bulbulodentata
├─Hyopsodontidae
└─Meridiungulata
├─Didolodontidae
│ ├─Didolodus
│ └─Litopterna
└─Notoungulata
├─Toxodontia
└─Typotheria
|
Contents
Overview |
Litopterans and LeprechaunsLet us confront the most critical issue first: is it really necessary to have big ears, a sharp nose, receding hairline and a little white beard to study South American ungulates? Since Florentino Ameghino 1854-1911), George Gaylord Simpson 1902-1984), and Richard Cifelli b. 1950-something) have overlapping lifespans, we are forced to discount reincarnation as a possible explanation. Yet the fact remains that, for well over a century, South American ungulate research has been spearheaded, often dominated, by people who look very much like middle-aged leprechauns. Dozo (1996) has observed that Miocene typotheres left brain impressions on the insides of their skulls which closely match those of the unrelated caviid rodents who lived at the same time and place, and shared a similar ecological niche. Could something of the sort be occurring on the outside of the skull as well, actuated merely by protracted puttering with Patagonian protocones? Note that notoungulates, too, have sharp rostra, inflated auditory bullae, and sagittal crests. This is a trend with alarming implications, particularly for those who study tapinocephalids easily the world's ugliest fossil vertebrates) or pareiasaurs....
These historical matters are not without some scientific relevance as well since, as is so often the case, we need to understand the Nineteenth Century view of things in order to see where we are today. Long before plate tectonics were understood, or even suspected, it became clear that South America had been faunally isolated from the end of the Mesozoic until just a few million years ago. This became quite clear in the closing decades of the Nineteenth Century, when Ameghino discovered and named innumerable ungulate species which were weirdly different from the better known ungulate groups of Europe, Africa, Asia and North America. Ameghino was a careful observer, a meticulous collector, and a brilliant descriptive scientist. Unfortunately, he was also something of a nut case on certain topics. Perhaps influenced by the extreme Latin nationalism of the times, Ameghino insisted that his South American fossils were the font from which most of the world's mammals, including man, had evolved. Perhaps this was a harmless eccentricity. Perhaps not. In any case, as other Gondwanan scientists have learned, the Laurasian scientific community tends to overreact to claims for Gondwanan primacy. Ameghino's anthropological work became something of a scientific scandal, and his paleontological work was badly discredited as an unfortunate side effect.
It was Simpson who did much to revive Ameghino's work in particular, and interest in South American ungulates more generally. As Simpson noted, Ameghino's descriptive work was head and shoulders above the standards of most Nineteenth Century paleontology. His theoretical errors, for that matter, were no worse than those of his contemporaries. The real problem, to Simpson, was that Ameghino was an over-splitter. When Ameghino's innumerable tooth taxa were distilled down to a workable number, and if one applied reasonably modern notions of evolutionary descent, the whole thing fell into a pattern not very different from what we recognize even today. This analysis left Simpson with the interesting issue of the South American groups as alternate ungulates. That is, he supposed the Latin fauna to be one gigantic monophyletic radiation of a single dispersal of primitive North American forms into an ecological vacuum, isolated from the rest of the world -- almost the converse of Ameghino's theoretical ideas.
Cifelli has, over the last 25 years or so, worked to test Simpson's vision rigorously. His objective has been to define precisely the relationships of the didolodonts and litopterans and their origin in the North American fauna of the Latest Cretaceous and Early Paleocene. The interrelationships at the South American end are complex and still a bit unclear. However, it is reasonably certain that the Didolodontidae are paraphyletic and include all of the Litopterna. Recently, Cifelli and Christian deMuizon [dC00] have published what appears to be a definitive paper establishing the origin of this group in the Kolpaniinae, a particular subfamily of North American "condylarths." Thus, as to this group, Simpson is vindicated. It is monophyletic, and its origins are in North America. But what of the other South American ungulates, particularly the largest and most diverse group, the Notoungulata? DeMuizon & Cifelli reach no firm conclusion, and Cifelli's opinion on the subject seems to have varied over the years. We will take this matter up on a succeeding page. ATW030105.
Added Note: Gheerbrant et al. (2001) have made a rather good dental case that the South American endemic ungulates may have come from North Africa, rather than North America. In either case, the method of dispersal is not easy to imagine. However, we are necessarily looking at a unique dispersal event which presumably occurred by some wildly unlikely combination of circumstances. ATW060321.
 Didolodontidae: Indalecia, Paulacoutoia, Victolemoinea
Didolodontidae: Indalecia, Paulacoutoia, Victolemoinea
Range: Early? Paleocene Thanetian) to Pleistocene of South America
Phylogeny: Meridiungulata : Notoungulata + * : Didolodus + Litopterna.
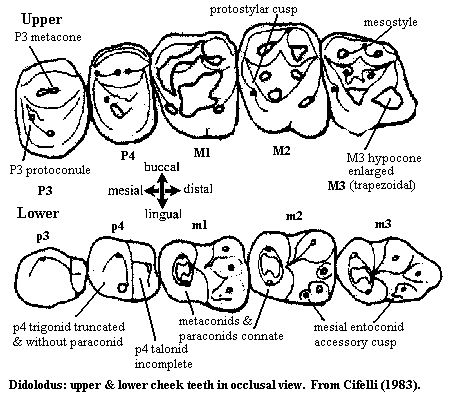
Characters: small-bodied link between "condylarths" and litopterns; distal upper premolars large and inflated [C83]; P3 primitively not strongly molariform [C83]; P4 with enlarged protocone [C83]; P3 protocone large and aligned lingual to paracone [C88]; P4 quadrate, with protocone & paracone similar height & volume [dC00]; p4 with cuspate talonid [$C93]; p4 with metaconid distal and almost equal to protocone [dC00]; labial cingulum usually present [dC00]; no cristae joining meta- & paracones with their corresponding conules (i.e. no preparaconular or postmetaconular cristae) [dC00]; hypocone on M1-2 [$C93]; M1-2 with parastylar lobe absent [dC00]; hypocone on M3 [C83] [$C93*]; primitively, upper molars without mesostyle [C83]; derived forms with mesostyle [S48]; talonid at least as wide as trigonid [dC00]; paraconid appressed to metaconid [$C93]; highly arcuate paracristid [dC00]; paraconids connate but distinct from metaconids [C83]; talonid basin open, but hypoconulid & entoconid well differentiated [dC00]; hypoconid usually larger than protoconid [dC00]; hypoconid invades talonid basin mesial to hypoconulid [dC00]; hypoconulid distinct from entoconid [dC00]; entoconid usually larger than hypoconulid [dC00]; cristid obliqua reaches distolingual base of protoconid or center of protocristid [dC00]; astragalus tibial trochlea more convex & better defined than in condylarths [d+98]; astragalus without sharp crests on margins of tibial trochlea [C83]; astragalar proximal foramen absent [C83] [S48]; astragalus medial face with strong anterior flare [$C93*]; astragalar facet for medial malleolus of tibia vertical & turns sharply medial anteriorly(?!) [C83] & well-developed [d+98]; astragalus sustentacular facet short and separate from ectal facet [S48]; sustentacular facet reaches or almost reaches articular surface of head [dC98]; astragalus ectal facet wide, oblique & deep [S48]; astragalus neck short, oblique and somewhat flattened dorsoplantarly [S48]; astragalus with deep body [C83] [d+98] (appears to contradict Simpson); astragalus with no medial projection, but with lateral projection [S48]; astragalar head medially placed [$C93*] [d+98] & transversely expanded [C83] [S48]; astragalus does not contact cuboid; convex navicular facet [S48]; fibular tubercle of calcaneum located dorsally [$C93*]; fibular tubercle of calcaneum extends posterodorsally & terminates at the base of the ectal prominence [C83]; calcaneum fibular facet tightly curved & trochlea-like [d+98]; .
Notes: [1] It has been clear for many years that the didolodontids are paraphyletic and include all of the litopterans [S48]. Thus it might be better to adopt the designation Didolodontoidea as suggested by Cifelli [C83], or just include both under Litopterna. The latter seems the better course. Not only are the didolodonts paraphyletic, but genera assigned to the didolodonts are "nearly indistinguishable" from primitive litopterns [C85]. Indeed, Cifelli has argued that some litopterns, the Sparnotheriodontidae, are more closely related to Didolodus than to derived litopterns [C93]. However, this clade is not terribly well supported, and was not found in his most recent analysis [dC00]. [2] synapomorphies with "[$C93]" are those found to unite the entire didolodontid + litoptern group. Synapomorphies with the asterisk "[$C93*]" relate to the Didolodontidae, sensu stricto. [3] as noted by [dC00], the enlarged cones turn the valleys between the protoconid and the other main trigonid cusps into two mesiodistally-oriented folds.
Links: Nathis Fauna Zuid-Amerikaanse hoefdieren Dutch; brief, non-technical description); pal3 species list); mammals2 basic range data); Untitled Czech); Paleocene mammals of the world brief discussion of ecology & evolution).
References: Carroll (1988) [C88]; Cifelli (1983) [C83]; Cifelli (1985) [C85]; Cifelli 1993) [C93]; deMuizon & Cifelli (2000) [dC00]; deMuizon et al. (1998) [d+98]; Simpson (1948) [S48]. ATW021226.
 Didolodus: Ameghino
1897: D. multicuspis Ameghino 1897. D. minor Simpson 1948. The best known didolodont genus.
Didolodus: Ameghino
1897: D. multicuspis Ameghino 1897. D. minor Simpson 1948. The best known didolodont genus.
Range: Late Eocene of Argentina.
Phylogeny: Didolodontidae : Litopterna + *.
Characters: 60 cm; premaxilla probably very short [S48]; doubled infraorbital foramen above P4 [S48]; root of zygomatic arch above M2-M3, sloping rapidly back & not diverging from line of maxilla [S48]; jugal covers lateral surface of zygomatic arch and reaches lacrimal [S48]; mandibular symphysis narrow and "ends opposite" P2 [S48]; jaw shallow & "plump" [S48]; dental formula ?/3, 1/1, 4/4, 3/3 [S48]; unworn enamel surfaces have fine wrinkles [S48]; no diastema [S48]; lower incisors almost transversely placed and procumbent [S48]; c1 alveolus twice size of incisor alveoli & only slightly procumbent [S48]; P1 roots divided at tip only [S48]; P1 with tall mesiobuccal cusp [S48]; P1 with small buccodistal accessory cusp [S48]; P1 with distal basined heel marked off by strong distal cingulum [S48]; premolars 3 & 4 of approximately same size & structure [S48]; P3 enlarged and about the same size as P4 [C83]; P3-4 transversely wide, with quadrate buccal corners [S48]; P3 metacone present [C83]; P4 metaconule and protoconule present [C83] [S48]; P3 & P4 paracone & metacone confluent or slightly separated [S48]; P3-4 protocones lower than buccal cusps [S48]; P3-4 cingulum, lingually weak or absent & with small para- & metastyles on mid-buccal side [S48]; p1 single-rooted & not procumbent [S48]; p2 with single central cusp and transversely wide basin distally bordered by crests & with one buccodistal?) cuspule [S48]; p3 poorly known, but probably like p2 [S48]; p3 with small metaconid [dC00]; p4 trigonid truncated, with paraconid reduced or absent [C83] (contra [S48] three cusps on trigonid); p4 without medial talonid cusp [C83] (?contra [S48], crest extending distally with cusp in middle of talonid); p4 with metaconid lingual and almost equal to protocone [dC00]; crests extend lingually & buccally from talonid cusp, so that talonid has two small walled basins [S48]; molars do not become sequentially larger distally [C83]; uppers with hypocone about same size as protocone [S48]; postparaconular & premetaconular cristae usually absent [dC00]; M1 quadrate and axially longer than premolars [S48]; M1 with trigonid cones of about same height & hypocone slightly lower [S48]; M1 protocone & hypocone compressed linguobuccally and drawn out into distinct crescents [S48]; M1 conules very sharp & distinct, about 50% size of main cusps [S48]; M1 conules slightly marginal to straight line between main cusps [S48]; M1 hypocone further from margin than protocone [S48]; M1 with strong mesostyle slanting buccodistally [S48]; M3 trapezoidal, with enlarged hypocone [C83]; M3 & m3 elongate [S48]; M3 enlarged [dC00]; M2 very similar to M1 [S48]; M2 buccal border less straight [S48]; M1 & M2 with buccal cingulum stronger than distal cingulum [S48]; uppers with continuous relatively broad cingulum [S48]; hypocone progressively more pronounced on distal molars [S48]; upper molars with weak but distinct protostylar cusp [S48]; upper molars with strong mesostyle [C83] [S48]; lower trigonids & talonids of about same size [S48]; lower molar paraconids connate but distinct from metaconid [C83] (contra fused [S48], almost indistinguishable [dC00]); trigonid basin closed by mesial & distal cristids from protoconid to fused meta/paraconid [S48]; talonids with large crescentic entoconid (??), smaller entoconid & very small hypoconulid distal to other two cusps [S48] (contra: entoconid small than hypoconulid [dC00]); cristid from metaconid to hypoconid with small cusp near midline [S48] [1]; lower molars with anterior entoconid accessory cusp [C83] [S48]; lowers with narrow precingulid [S48]; m3 with talonid transversely?) narrower than trigonid & larger hypoconulid [S48].
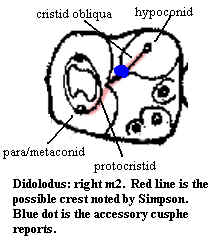 Notes: [1] most (not all) of the differences between the descriptions of Simpson and Cifelli can be explained by Simpson's less precise terminology. This one is tougher. See image at right. The crest referred to could be the cristid obliqua which seems to merge with the protocristid so as to create the impression of a single cristid. However Cifelli does not note an accessory cuspid here. Simpson cannot mean the entoconulid ("entoconid accessory cusp" of Cifelli), since Simpson notes this separately. Perhaps he is seeing a cusp (or Cifelli is missing one) based on wear at the rather complex intersection of the hypoconid, cristid obliqua, protocristid & trigonid/talonid interface.
Notes: [1] most (not all) of the differences between the descriptions of Simpson and Cifelli can be explained by Simpson's less precise terminology. This one is tougher. See image at right. The crest referred to could be the cristid obliqua which seems to merge with the protocristid so as to create the impression of a single cristid. However Cifelli does not note an accessory cuspid here. Simpson cannot mean the entoconulid ("entoconid accessory cusp" of Cifelli), since Simpson notes this separately. Perhaps he is seeing a cusp (or Cifelli is missing one) based on wear at the rather complex intersection of the hypoconid, cristid obliqua, protocristid & trigonid/talonid interface.
Links: New Page 1 vague life reconstruction); Species Skull Ht Lt Wt Time Range Acama probably Acamana U ... Range data); Paleontologia de Argentina - Eoceno (Spanish); EOCENO small image of teeth); Critter Gallery interesting life reconstruction).
References: Cifelli (1983) [C83] deMuizon & Cifelli 2000) [dC00]; Simpson (1948) [S48]. ATW040116.
Range: Early Paleocene to Pleistocene of South America.
Phylogeny: Didolodontidae : Didolodus + *.
Characters: horse & camel-like forms; tympanic not inflated, no epitympanic sinus & no ossified auditory meatus [S48]; dentary symphysis fused in derived forms [dC00]; dental formula variable: 1-3/2-3, 0-1/1, 4/4, 3/3 [S48]; teeth generally low-crowned and lophodont [C88]; one pair of incisors may be enlarged and rootless, otherwise incisors similar size [S48]; canines small & undifferentiated [S48] [dC00]; premolars generally have a swollen appearance; primitively, P1 & p1 single-rooted; distal premolars molariform [S48]; distal upper premolars large and inflated [C83]; P3 with broadly sweeping postprotocrista which forms distinctive distal platform; P4 4-sided (parallelogram?); primitively, p4 somewhat molarized, with metaconid as large as protoconid and often distal to it [dC00]; P4 with reduced styles (=stylar cusps?) & strong pre- & postcingula; in derived forms [C83]; p4 with metaconid lingual and almost equal to protocone [dC00]; upper molars quadrate with crescentic paracones & metacones [C83] [S48]; M1-2 with parastylar lobe absent [dC00]; derived forms with para- and mesostyles on upper molars [C83]; protoloph imperfect without full union of cusps [S48]; metaloph absent [S48]; crochet absent [S48]; M3 usually large; primitively, labial cingulum present but incomplete on upper molars [dC00]; primitively, paracone larger & taller than metacone; hypocone present, but much smaller than protocone [S48]; primitively, postparaconular & premetaconular cristae absent [dC00]; parastyle reduced or absent; in derived forms, lower molars bicrescentic [C83] [S48]; derived forms with cristid obliqua attaching to base of metaconid mesially [C83] (contra cristid obliqua reaches distolingual base of protoconid or center of protocristid [dC00]); primitively, protocristid oblique, with distally concave [dC00]; primitively, m3 unreduced in size; m3 without third (hypoconulid?) lobe [S48]; primitively, trigonid not wider than talonid and only moderately taller [S48]; talonid basin open, but hypoconulid & entoconid well differentiated [dC00]; primitively, metaconid well distal to protoconid & expanded distally [dC00]; paraconid varies widely, from very large to absent (contra [S48]: paraconid distinct from and same size as metaconid); primitively, paraconid more lingual than buccal; highly arcuate paracristid [dC00]; primitively, hypoconulid usually smaller than entoconid and hypoconid larger than protocone; anterior end of hypoconid crescent at or near metaconid [S48]; primitively, entoconid placed distally & connate with hypoconulid [S48?]; paracristid narrower than half width of trigonid, deeply arched posteriorly, projected anteriorly, & attaching to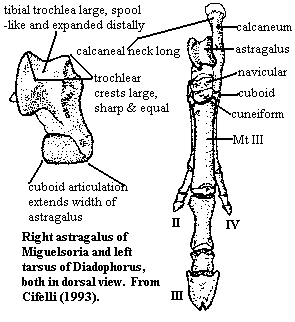 anterolingual side of proto- & paraconids; primitively, strong precristid present; entocristid absent; limbs short [C88]; carpus with enlarged magnum and medial displacement of distal elements so that lunate does not contact unciform, but magnum contacts cuneiform [C83] [$C93]; manus/pes tridactyl but functionally monodactyl (i.e. mesaxonic) [C83] [$C93] [S48]; astragalus body spool-like & without proximal astragalar foramen [C83]; tibial trochlea deep, with elevated crests [S48]; astragalus tibial trochlea extending posteriorly (?) & distally from dorsal surface [$C93]; astragalus with prominent equal tibial & fibular crests [C83] [$C93] [S48]; astragalus sustentacular facet "anteroposteriorly elongated" (?) [C83]; astragalus ectal facet deeply concave "with pronounced outward orientation" (= friendly?) [C83] [$C93]; astragalar neck moderate length & slightly constricted [S48]; astragalar head roughly semicylindrical [C83]; astragalar head wide [S48]; navicular facet extending onto both sides of astragalar head [C83] [$C93]; astragalus medial collateral ligament facet absent [$C93]; astragalus cuboid facet absent [$C93] [S48]; calcaneum neck long with "strongly oblique cuboid facet 1" [$C93]; necks of proximal tarsals primitively elongated [C83]; earlier forms rabbit-sized & specialized for jumping, with larger, late forms very horse-like & adapted for fast running [C85].
anterolingual side of proto- & paraconids; primitively, strong precristid present; entocristid absent; limbs short [C88]; carpus with enlarged magnum and medial displacement of distal elements so that lunate does not contact unciform, but magnum contacts cuneiform [C83] [$C93]; manus/pes tridactyl but functionally monodactyl (i.e. mesaxonic) [C83] [$C93] [S48]; astragalus body spool-like & without proximal astragalar foramen [C83]; tibial trochlea deep, with elevated crests [S48]; astragalus tibial trochlea extending posteriorly (?) & distally from dorsal surface [$C93]; astragalus with prominent equal tibial & fibular crests [C83] [$C93] [S48]; astragalus sustentacular facet "anteroposteriorly elongated" (?) [C83]; astragalus ectal facet deeply concave "with pronounced outward orientation" (= friendly?) [C83] [$C93]; astragalar neck moderate length & slightly constricted [S48]; astragalar head roughly semicylindrical [C83]; astragalar head wide [S48]; navicular facet extending onto both sides of astragalar head [C83] [$C93]; astragalus medial collateral ligament facet absent [$C93]; astragalus cuboid facet absent [$C93] [S48]; calcaneum neck long with "strongly oblique cuboid facet 1" [$C93]; necks of proximal tarsals primitively elongated [C83]; earlier forms rabbit-sized & specialized for jumping, with larger, late forms very horse-like & adapted for fast running [C85].
Note: primitively, as noted by [dC00], the enlarged cones turn the valleys between the protoconid and the other main trigonid cusps into two mesiodistally- oriented folds.
Image: Macrauchenia.
Links: Litopterna; Untitled (Czech); de Muizon & Cifelli; Estudios Geológicos, vol.53 (5-6), 1997 Spanish); The Journal of Vertebrate Paleontology; Paleocene mammals of the world; Paleontology and Geology Glossary- Ma.
References: Carroll (1988) [C88]; Cifelli (1983) [C83]; Cifelli (1985) [C85]; Cifelli 1993) [C93]; deMuizon & Cifelli (2000) [dC00]; Simpson (1948) [S48]. ATW021224.
checked ATW040708
