 Learning by Ear
Learning by Ear| Meridiungulata | ||
| The Vertebrates | Notoungulata |
| Vertebrates Home | Vertebrate | Vertebrate |
|
Abbreviated Dendrogram
Laurasiatheria
│
└─Meridiungulata
├─Litopterna
└─Notoungulata
├─Henricosborniidae
└─┬─Notostylopidae
└─┬─Toxodontia
└─Typotheria
|
Contents
Overview |
The Notoungulata, as currently understood, currently include 13 families and over 100 genera. [C85]. They are certainly an early group. A diagnostic notoungulate molar is known from the Early Paleocene. [dC00]. All of the basic groups had differentiated by Late Paleocene [C85]. During the Miocene, they underwent another pulse of change, perhaps driven by the increasing aridity of South America and the arrival of rodents. However, their overall diversity probably never recovered to Eocene levels. Their numbers collapsed during the Pliocene, and most were extinct before a permanent land bridge with North America developed in the Middle Pliocene, bringing with it a broad invasion by North American mammals of all kinds. [C85].
The most basal of the Notoungulata have traditionally been placed in the "Notioprogonia." As Simpson stated long ago, this is not a natural group [S48], and we will accordingly avoid referring to it. Two families of early notoungulates do seem to be distinctive: the Henricosborniidae and Notostylopidae. These two families were probably extinct before the end of the Eocene. An isolated report of a Miocene survival [M56] appears to have been refuted [CV97]. These basal forms were succeeded by the large pig and hippo-like Toxodonta and the rabbit and rodent-like Typotheria.
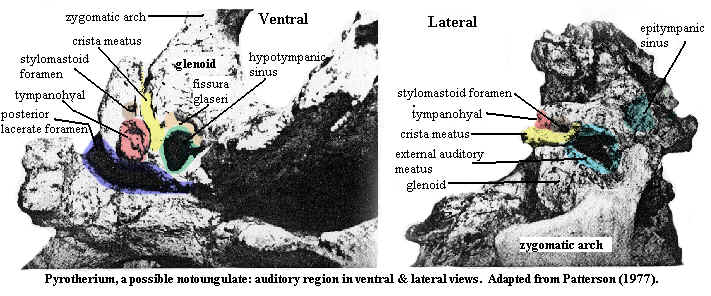
Learning by EarIn addition to the usual arcana of dental detail, notoungulates are characterized by specializations of the auditory area of the skull. The notoungulates possessed strongly inflated auditory bullae. Numerous literature sources also dutifully recite that the notoungulates were also characterized by: "canal of Huguier opening at the posterior end of the fissura Glaseri," "large vagina of the processus hyoidei," and "crista meatus" -- as if these terms were known to anyone with an elementary school education. In point of fact, some of these phrases are obsolete and survive only by rote repetition in the paleo literature. We will rise to this challenge and try with help from Patterson [P77]) to explain what is actually going on.
The best place to start may be with the n. chorda tympani, a nerve which carries taste sensation from the mouth, among other things. Beginning at the brain end of things, the chorda tympani is initially part of the facial nerve. The facial nerve exits at the stylomastoid foramen, the brown spot lateral to the big red tympanic complex in the figure. However, the chorda never gets there. Instead, it branches from the facial nerve and wanders off through a tiny canal right into the middle of the middle ear. Why evolution permits a critical taste nerve to cut across an equally critical auditory tympanic membrane is difficult to imagine, but this is a consistent feature of mammalian anatomy. The nerve is called the chorda tympani precisely because it cuts a chord across the surface of the tympanum. Now, having wandered into the acoustic equivalent of the Gare du Nord, it naturally seeks to leave expeditiously. It does so by another little canal, the anterior chordal canal which, 150 years ago, went by the name of canal of Huguier (often misspelled Hugier). This anterior chordal canal then opens into a larger space, the petrotympanic fissure between the bulla and the basicranium, through which it completes its getaway. As you may have guessed, the petrotympanic fissure was once called the fissura Glaseri. In most mammals, the canal leads into the fissure in the middle. In notoungulates, the canal joins the fissure at its posterior end. That's all this means.
OK, so what's this "large vagina of the processus hyoidei"? The tympanic membrane is relatively delicate and requires more structural support than the thin ring of bone which suspends it. The bone immediately adjacent to the tympanum may get built up, both as physical support internally) but also to provide one more mechanism for focusing sound on the membrane (externally). This is the vaginal process. Notoungulates have an ossified outer ear (external auditory meatus), like some primates, so it is not surprising that they also have a large vaginal process around the tympanic membrane.
Two other aspects of the notoungulate ear are the crista meatus surely they mean crista meati!?) and the various accessory tympanic sinuses. Here, note that our view of the tympanohyal in the left-hand image is, essentially, from the inside of the middle ear. The opening of the outer ear is shown in the right-hand image. The external auditory meatus runs as a bony tube to the side of the tympanohyal we cannot see on the left-hand picture. This tube has a large ridge, more or less ventral to the meatus. This is the crista (yellow in the image). The various ancillary sinuses (epitympanic, hypotympanic, and so on) are just that: empty places in the skull connected in some fashion to the middle ear. Their functional significance is unknown.
While we have done our best to explain this difficult region, we admit to considerable doubt about our success in doing so. Fortunately, a far better electronic resource has recently become available, since this essay was originally posted, to which the reader is enthusiastically referred. Gabbert 2004), which may be found on the web at this page of the AMNH site.
 Meridiungulata or Panameriungulata? -- The Lophed World of the Notoungulata
Meridiungulata or Panameriungulata? -- The Lophed World of the NotoungulataAs discussed on the meridiungulate page, there is no consensus about the relationships of the Didolodontidae and the notoungulates. DeMuizon & Cifelli [dC00] place a box called Panameriungulata around the didolodonts, litopterns and certain Laurasian "condylarths," but they can't tell us whether the notoungulates are in the box. The reason for the difficulty is teeth. Although we have reasonably good ears -- and also some ankles -- from notoungulates, these are mostly Miocene or later. These are far to late to tell us much about notoungulate origins. We must rely on teeth, and there the notoungulates confront us with a major difficulty, for they have lophed their teeth. That is, instead of the relatively orderly pattern of cusps possessed by most mammals, notoungulates have drawn out the cusps into long, messy arcs. The general pattern is shown in the image at right from Carroll [C88]. Thus comparison is quite difficult.
Nevertheless, we will part company with deMuizon & Cifelli's careful and scholarly work on this point. Or, rather, we will embrace their work but be far less careful and scholarly in applying their results. After all the dust has settled at the end of the paper, [dC00] summarize the characters which unite their Panameriungulata. Those characters include the following:
1. The metaconid is distally placed. This is a character plainly present in both basal families of Notoungulata (refer to the figures below under Henricosborniidae and Notostylopidae). The linguodistal corner of the trigonid is drawn out and the metalophid is consequently oblique. This is an especially marked character of the henricosborniids, as noted by Simpson [S48].
2. The entoconid is distally placed and connate with the hypoconulid. This character is not present in notoungulates. However, it is also absent in primitive didolodonts [dC00]. As these authors remark, this character is "very homoplastic" and "of problematic use in phylogenetic reconstruction" [dC00: 131]. So the absence of this trait tells us nothing..
3. Development of a lingual talonid notch. This character is not present in notoungulates, but seems to be entirely correlated with the previous character. The "notch" is caused by the distal displacement of the entoconid (which normally forms the lingual wall of the talonid basin) and its merger with the hypoconulid. Review of the dental diagrams of didolodonts and basal litopterns reveals that it is absent or very weak in these forms as well. Conversely, more derived notoungulates, like derived litopterns, possess a strong notch (see the Notostylops figure, below). We may speculate that this character is simply a result of scaling up, for a large animal, a basic pattern of dentition shared by the smaller ancestors of both notoungulates and litopterns.
4. Reduction or loss of the postparaconular and premetaconular cristae. This character is present in all notoungulates. In fact, notoungulates generally have no cristae, since they rely on prolongation of the cusps themselves into lophs. Thus the comparison is a little unfair.
So far, so good. But this is not very convincing, as it reveals only one valid synapomorphy. However deMuizon and Cifelli also discuss several additional synapomorphies uniting only the South American forms. These include:
5. The metaconid is displaced so far distally that it projects sometimes weakly) into the talonid basin. This appears to be present in notoungulates, although it is not strongly marked in this group or, for that matter, in didolodonts.
6. The paracristid is strongly arcuate, extending mesially from the protoconid, making a sharp turn lingually, and then turning distally to reach the paraconid. We must be cautious here for two reasons. First this is a character which is quite unique in mammals, a strong synapomorphy of the South American branch of the panameriungulates. Second, notoungulates have few conventional cristae, no paraconid as such (a trait shared with some didolodonts), and often a neomorphic cusp in the middle of the trigonid. Given these differences, the actual resemblance is striking. Notoungulates do possess a crest (loph or crista?) running straight mesially from the protoconid to the mesiobuccal corner of the molar, where it makes the same right-angle turn down the mesial face of the tooth. Since there is no paraconid, we cannot make the final comparison, but there certainly is a crest of some sort that runs distally from the mesial edge in many of the lower molars figured by Simpson [S48] for this group. Thus, we can be reasonably confident that this critical synapomorphy is shared by notoungulates and endemic South American panameriungulates.
7. The p4 metaconid is transversely aligned with the protoconid and about the same size. This trait is not present in notoungulates, all of which have a rather unique and specialized p4 trigonid. However, deMuizon & Cifelli note that this trait is "equivocal" because it is not shared by some of the earliest South American forms.
Ultimately, the evidence is not strong, but the most parsimonious conclusion is that notoungulates are members of the Panameriungulata, as defined by [dC00]. Based on character #7 above, it is likely that they diverged from the basal "condylarth" stock after, but just after, its initial arrival in South America in the latest Cretaceous or earliest Paleocene. Accordingly, we believe that the evidence continues to support Simpson's hypothesis of a monophyletic South American radiation combining both notoungulates and the didolodont - litoptern clade. ATW030105.
References: Carroll (1988) [C88]; Cifelli (1985) [C85]; Cifelli (1993) [C93]; Cifelli & Villarroel (1997) [CV97]; deMuizon & Cifelli (2000) [dC00]; McKenna 1956) [M56]; Patterson (1977) [P77]; Simpson 1948) [S48].
Range: Paleocene to Pleistocene of South America.
Phylogeny: Meridiungulata : Didolodontidae + * : Henricosborniidae + (Notostylopidae + (Toxodontia + Typotheria)).
Characters: skull generally small & pear-shaped [S48]; zygomatic arch strong & high on skull [S48]; posterior base of zygomatic arch originating high on side of skull [$C93]; external auditory meatus formed by ossified tube [C88] [$C93] [S48] ossified auditory meatus often with crest [S48], on ventral surface [P77]; inflated auditory bulla [P77]; epitympanic sinus developed in squamosal [$C93] [S48]; accessory chambers above (= epitympanic sinus?) and below auditory bulla [C88] [P77] [S48 -- states that they are venous sinuses]; hypotympanic sinus particularly large [P77]; additional venous sinus system associated with all this [P77]; stylomastoid foramen opens between crista on auditory meatus and post-tympanic portion of squamosal [P77]; mandibular symphysis fused [dC00]; primitively, canines reduced [dC00]; primitively, complete dentition without diastema [C88]; cheek teeth generally lophodont [S48]; premolars always somewhat molarized, but rarely to a great extent [S48]; P1 & p1 single-rooted; primitively, teeth low-crowned [C88]; upper molars with straight, prominent buccal ectoloph & oblique mesiolingual protoloph [C88] [S48]; upper molars with distolingual metaloph, often with anteriorly-directed crochet [C88] [$C93]; lower molars generally with short, asymmetrical trigonid crescent & longer, more symmetrical talonid crescent [S48]; lower molars with entoconid in transverse distolingual entolophid located inside curve of hypolophid [C88] [$C93] [S48]; paraconid absent [$C93]; paracristid short [$C93]; protoconid and metaconid united by metalophid [C88]; astragalus tibial trochlea primitively shallow and more dorsal than proximal [S48]; astragalus with medial protuberance, long, constricted neck & sulcus extending laterally from the superior astragalar foramen [$C93]; primitively 5 digits, but often 3-toed & mesaxonic in derived forms [S48]; hoofed unguals (some exceptions) [S48].
Image: (right) Huilatherium pluriplicatum skull in palatal view, from La Venta Fauna: Fossil Gallery. Huilatherium is a derived toxodont from the middle Miocene of Columbia, described in more detail in Villarroel & Danis (1997).
Links: Notoungulata Mikko); xrefer - Notoungulata; Untitled (Czech); pal4 lists); de Muizon & Cifelli; extinct ungulate page.
References: Carroll (1988) [C88]; Cifelli (1993) [C93]; deMuizon & Cifelli (2000) [dC00]; Patterson 1977) [P77]; Simpson 1948) [S48]. ATW021225.
 Henricosborniidae: Camargomendesia, Henricosbornia, Othnielmarshia, Peripantostylops, Simpsonotus.
Henricosborniidae: Camargomendesia, Henricosbornia, Othnielmarshia, Peripantostylops, Simpsonotus.
Range: Late Paleocene Ríochican [S48]) to Late Eocene (Early Casamayoran [C85]) of South America, with possible survival to the Late Oligocene Deseadan [C85]).
Phylogeny: Notoungulata : (Notostylopidae + (Toxodontia + Typotheria)) + *.
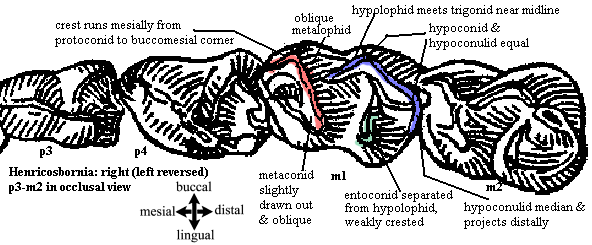
Characters: small sized [S48]; cheek teeth low-crowned [S48]; upper premolars triangular with simple protocone [S48]; premolar hypocone absent [S48]; P3-4 with distinct paracone & metacone [S48]; p3-4 molariform and simply bi-crested [S48]; cristae generally weak to absent [S48]; M1-2 with equal, distinct protocone & hypocone (more distinct than in notostylopids) [S48]; protoloph simple & oblique [S48]; protoconule vestigial [S48]; metaloph short & transverse [S48]; crochet on metaloph variable but generally short and not reaching ectoloph [S48]; crochet shorter & more oblique than notostylopids [S48]; ectoloph with distinct paracone & metacone [S48]; mesostyle vague or absent (like teenagers) [S48]; M3 triangular with very short metaloph, often weakly crested [S48]; trigonid crests (= metalophid?) oblique [S48]; metaconid taller than protoconid & slightly drawn out mesiobuccally - distolingually [S48]; median trigonid cusp absent (compare notostylopids) [S48] [1]; metalophid runs straight mesially from protoconid & makes right angle turn lingually at buccomesial corner of tooth [S48]; hypoconid & hypoconulid of similar size [S48]; hypoconulid projects distally [S48]; hypolophid meets trigonid (metalophid) near midline [S48]; entoconid, weakly crested, distinct from hypolophid, & positioned relatively distal [S48].
Notes: [1] but the m2, and perhaps m3, trigonids in the figure appear to have median cusps, and the m2 median cusp is quite large!
References: Cifelli (1985) [C85]; Simpson 1948) [S48]. ATW021225.
 Notostylopidae: Edvardotrouessartia, ?Homalostylops, Notostylops, Otronia, ?Seudenius
Notostylopidae: Edvardotrouessartia, ?Homalostylops, Notostylops, Otronia, ?Seudenius
Range: Late Paleocene Ríochican) to Late Eocene Mustersan) of South America. [M97]. Rare, perhaps absent, in Ríochican [S48].
Phylogeny: Notoungulata :: (Toxodontia + Typotheria) + *.
Characters: skull triangular (in dorsal view?) [S48]; rostrum short & narrow [S48]; nares terminal [S48]; skull table broad & low [S48]; usually strong sagittal & lambdoid crests [S48]; zygomatic arches quadrate [S48]; "strong postorbital constriction" [S48] [1]; epitympanic sinuses relatively small (i.e. small for a notoungulate) [S48]; hypotympanic sinus large & globular, extending well below basisphenoid [S48]; ossified external auditory meatus short and horizontal [S48]; dentary elongate, straight and slender [S48]; dental formula 2-3/2-3, 0-1/0-1, 4/4, 3/3 [S48]; large nipping, styliform incisors separated from cheek teeth by diastema [C85] [C88] [$C93]; 1st & 2nd incisors enlarged but rooted [S48]; 3rd incisors, canines & 1st premolars reduced & styliform or absent, leaving diastema [S48]; loss of some anterior dentition [$C93]; cheek teeth low-crowned, and more strongly lophodont than henricosborniids [S48]; P1-4 triangular to quadrate [S48]; p2-4 progressively more molariform, but with heel short & entoconid less separate from outer crest (means loph?) than on molars [S48]; cristae absent [S48]; M1-2 quadrate, wider than long [S48]; M1-2 with hypocone and protocone connate but separated in lingual face by groove(s) [S48]; crochet longer & less oblique than henricosborniids [S48]; protocone less well separated from hypocone than in henricosborniids [S48];  protoloph simple & not completely confluent with ectoloph [S48]; metaloph more confluent with protoloph & with long crochet running straight mesially & ending near protoloph [S48]; trigonids with median crest having three small cuspules [S48]; talonid with simple outer hypolophid & simple transverse ectoconid loph [S48]; hypolophid reaching protolophid near midline [S48].
protoloph simple & not completely confluent with ectoloph [S48]; metaloph more confluent with protoloph & with long crochet running straight mesially & ending near protoloph [S48]; trigonids with median crest having three small cuspules [S48]; talonid with simple outer hypolophid & simple transverse ectoconid loph [S48]; hypolophid reaching protolophid near midline [S48].
Notes: [1] How can this be if the skull is triangular?
Links: pal4; mammals2; Nathis Fauna Zuid-Amerikaanse hoefdieren.
References: Carroll (1988) [C88]; Cifelli (1985) [C85]; Cifelli 1993) [C93]; Maas 1997) [M97]; Simpson 1948) [S48]. ATW021225
checked ATW040708
