Euteleostei: Neognathi
Umbra & Dallia
Taxa on This Page
- Dallia
- Umbra
La Cathédrale Engloutie: Ecology and Esociformes

The Kingdom of Ys is the French version of Atlantis. It is an old Breton story with a hundred variations. Almost all of them involve a drowned city with a great cathedral. At long intervals, its towering spires may still rise above the waves and its ancient bells then toll across the waters. This was the aspect of the tale that Claude Debussy attempted to capture in his tone poem, La Cathédrale Engloutie. Our purposes are considerably more trivial, of course. Yet there is something compelling in the image of a complex tale lost in Laurasian waters for 70-80 million years. We can learn much by the usual digging and diving; but we may learn a bit more yet by watching and listening to what can still be observed today -- at least when the tide is right.
To return to from impressionism to ichthyology, I am suggesting that behavioral and ecological characters of living fishes may be useful to understanding the evolution of Esociformes. Although Esox and Umbra diverged back when the typical citizen of Canada was a hadrosaur, they still share a number of behavioral and ecological characters which deserve more study. We have no solutions to offer, but -- at a minimum -- a new set of characters which ought to be taken into consideration.
By way of background, let us begin with a little paleontology of the conventional sort. Esocids, namely Estesesox, Oldmanesox and unnamed bits and pieces, are known in North America from the Late Cretaceous Campanian). Wilson et al. (1992). These are tiny creatures, the size of Umbra today, but recognizably members of Esocidae. They are known only from isolated dentaries and palatines, so we have little idea of their overall form. By the Early Paleocene, the modern pike genus, Esox is known from very complete specimens in western Canada. Wilson (1980); Wilson (1984). The Paleocene model, E. tiemani, is still on the small side, about the size of Dallia at 29 cm. However, it is morphologically almost identical to modern pikes, save for a considerably less expanded "bill" and some peculiarities of opercular morphology. Wilson (1984).
Since esocids were present in these high and far-off times, we may surmise that Umbra-like fishes were also lurking about somewhere. However, there is no direct evidence of them until the Oligocene. At that point we have Palaeoesox, which may be a big Umbra-like form, or a representative of some related group. Less directly, but more convincingly, the extant Umbra krameri is known from the Danube watershed, while all other Umbra are from North America. As there are no marine esociforms known, much less marine members of the genus Umbra, we are -- "forced" would be too strong a word -- but pushed urgently towards the conclusion that this genus diverged no later than the Eocene. This is the last possible time that a direct freshwater transfer was possible between Europe and North America. [1]
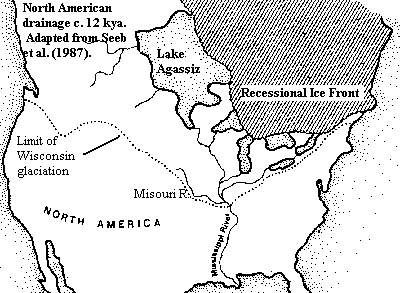 Thus it may seem that the two largest genera of esociforms were in place by the early Cenozoic and lived happily ever after. However that is not the usual interpretation. The distribution of esociform species is disjunct. That is, they tend to be found in ranges which do not overlap and which are separated by areas with no closely related species. This is most obvious in the case of Umbra, as one species inhabits Southeastern Europe and two others are found only in the eastern and central United States (and contiguous Canada). No Umbra inhabit western North America or Asia, both historically important areas of esociform diversity.
Thus it may seem that the two largest genera of esociforms were in place by the early Cenozoic and lived happily ever after. However that is not the usual interpretation. The distribution of esociform species is disjunct. That is, they tend to be found in ranges which do not overlap and which are separated by areas with no closely related species. This is most obvious in the case of Umbra, as one species inhabits Southeastern Europe and two others are found only in the eastern and central United States (and contiguous Canada). No Umbra inhabit western North America or Asia, both historically important areas of esociform diversity.
The interpretation usually given these facts is that we are observing a rebound of relict populations, and that the bulk of esociform diversity and numbers was destroyed by the Ice Ages of the Pleistocene. With respect to Esox lucius, the only species for which we have a good North American fossil record, both genetics and paleontology suggest that the entire continent was repopulated after the last (Wisconsin) glaciation from two or three small refugia with quite limited diversity. Crossman (1996); Seeb et al. (1987).
The significance of these historical events is that we should not rely blindly on cold adaptations to infer relationships. All esociforms are quite tolerant of low temperatures. Umbra, in particular, can maintain remarkably high metabolic levels close to 0° C. and is said to be able to withstand at least short periods of freezing. Chilton et al. (1984). We cannot assume that these characteristics are primitive, because of the harsh selection for cold adaptations in the Pleistocene, long after the living esociform genera had already diverged.
However, the esociforms share a number of ecological features which are less easily explained. These include:
a) Close association with vegetation. It would be difficult to overemphasize this similarity. All esociforms are restricted to heavily vegetated areas, both as fingerlings and adults. Bry (1996); Craig (1996); Page & Burr (1991). Esox (and perhaps others genera) has evolved an adhesive organ by which larvae adhere to vegetation as a normal part of the life cycle. As adults, Esox adopts an unusual hunting strategy which involves ambush predation from the edge of the vegetated zone into more open water. Bry (1996). All esociforms lack fin spines, which appears to be a distinct adaptation of fishes who spend their lives in heavy vegetation. Craig (1996).
 b) Visual predation: All esociforms have weak or absent lateral line organs and rely primarily on sight for predation. Accordingly, we may speculate that even the small members of the clade must prefer a habitat near the edge of the vegetated zone, with adequate light. Certainly, all members of the group are found in relatively shallow waters, and all have a low tolerance for turbidity. Bry (1996); Craig (1996); Page & Burr (1991).
b) Visual predation: All esociforms have weak or absent lateral line organs and rely primarily on sight for predation. Accordingly, we may speculate that even the small members of the clade must prefer a habitat near the edge of the vegetated zone, with adequate light. Certainly, all members of the group are found in relatively shallow waters, and all have a low tolerance for turbidity. Bry (1996); Craig (1996); Page & Burr (1991).
c) Low energy waters: All esociforms prefer low-energy environments. In part, this may be explained by the fact that esociforms are generally poor swimmers. The common description of pike as "sprint predators" (e.g. Crossman (1996)) may leave the impression that they are quick over short distance. In fact, Esox almost always attacks within a single body length. Hunting success for targets further away drops off very rapidly, and the pike loses virtually any chase. Webb & Skasden 1980). Nevertheless, there are reports of seasonal migration of Dallia we are not sure how well-established this behavior is), and occasional long-distance movement by certain individuals of Esox. Craig (1996). Umbra lacks the extreme posterior location of the dorsal & anal fins which restricts the distance swimming ability of the other esociforms. Nevertheless, it likewise avoids strong currents.
d) Tolerant of low oxygen: All esociforms can survive in low-oxygen waters and are probably capable of breathing air. Umbra can reportedly survive significant periods out of water. Esox and Umbra have well developed swim bladders adapted for air-breathing. Dallia, oddly enough, has adapted its esophagus to perform the same function. Crawford 1974).
e) Cannibalism. Esox and Umbra are both notable cannibals. We lack information on Dallia and Novumbra. At least in Esox, this may be part of an adaptation to ensure population survival. Pike grow at rates which are quite variable. Some smaller members of the year class (less than 200g) are typically eaten by larger pike at the end of the summer or as other food stocks decline. However, slower-growing pike are also more likely to survive particularly long or cold winters, presumably due to lower metabolic requirements. Craig (1996).
Reading over this list, it is clear that we are dealing with fishes strongly adapted to a fairly narrow environmental zone. Since Umbra and Esox diverged at least 50 Mya, and probably nearer 100 Mya, this is fairly significant stability. The basic ecological and behavioral parameters of the esociforms have been substantially more stable and striking, than, for example, the pattern of intramuscular bones, the caudal skeleton, the shape of the operculum, and presence or absence of nasals or extrascapulars -- all of which are used by workers to infer phylogeny. It may seem odd, given that the field is in full retreat from the alleged "fuzziness" of morphological characters, to promote the use of yet "fuzzier" behavioral and ecological characters. However, things are not always what they seem, and nothing tells this story as well as the esociforms. Molecular studies of primary structure are sometimes misleading for reasons which are not intuitively obvious. The ecological facts, by contrast, are known to every angler, and turn out to be quite stable and potentially useful.
Ecological characters have other advantages as well. Unlike isolated nucleotide or amino acid substitutions, ecological characters speak directly to the conditions of speciation, the role of adaptation, the kinds of environments experienced by a taxon, and its place within the community. In such cases, we can mark the precise spot where the genetic hammer strikes the species against the anvil of its environment. So, if we are fortunate -- if the tide and wind are right -- we may still hear the bells of the drowned cathedral and perceive at first hand the evolutionary events in world much older and deeper than Ys. ATW030425.
Image: (left) Northern pike, Esox lucius, by John F. Scarola, formerly posted at Discover the Fishes of All of Canada.
[1] Maybe. The present distribution of Dallia suggests that transfers via Asia might, conceivably, have occurred in much more recent times. No Umbra, alive or dead, are known from anywhere in Asia at any time. However, given the poor fossil record of small, freshwater teleosts, this negative evidence is not enough to eliminate the possibility.
Descriptions
 Umbra: Kramer 1777. U. krameri Walbaum 1792; U. limi Kirtland 1841; U. pygmaea DeKay 1842.
Umbra: Kramer 1777. U. krameri Walbaum 1792; U. limi Kirtland 1841; U. pygmaea DeKay 1842.
Range: U. krameri is found in the Danube drainage in SE Europe and appears to be primitive. The other species are from central and eastern North America, respectively.
Phylogeny: Esociformes : Dallia + (Novumbra + Esocidae)) + *.
Characters: maximum size 10-15 cm, depending on species; head bluntly conic; snout short, equal to diameter of eye; mouth moderate; premaxilla borders ~1/4th of gape [WV82]; ![U. limi left suspensorium from [WV82]](images/UmbraSuspensorium.gif) posterior premaxillae not protractile; premaxilla without dorsal articular process [WV82]; maxilla deepest posteriorly [WV82]; supramaxilla absent [WV82]; frontals terminate anteriorly below posterior 1/3rd of proethmoid [WV82]; frontal comes to a point over sphenotics [WV82]; frontal transverse ridge absent [WV82]; frontals overlie variable?) portion of supraoccipital [WV82]; parietals small (intermediate between Novumbra and Dallia) [WV82]; posttemporal small [WV82]; suspensorium short & deep [WV82]; symplectic not straight [W84]; preopercular with ventral arm larger than dorsal [WV82]; opercular with small lateral process, stout reinforcing rays, and slight posterior excavation [WV82]; subopercular & interopercular both triangular, of equal size and lack processes [WV82]; basioccipital with anteriorly-directed median spur (unique to Umbra) [WV82]; exoccipitals extended in posterodorsal- anteroventral axis [WV82]; sphenotic with lateral process at apex for origin of m. levator arcus palatini [WV82]; alisphenoids do not meet on the midline [WV82]; alisphenoid with large, ventrally- directed flange [WV82]; proethmoid anterior margin partially endochondral, curved ventrolaterally [WV82]; proethmoid anterior margin expanded & articulates with dorsolateral surface of premaxilla [WV82]; proethmoid thickened & grooved posteriorly to cap anterior end of rostral cartilage & articulate via cartilaginous joint with anteromedial margin of palatines [WV82]; vomer small, generally antero-posteriorly oriented & located variably ventral or under parasphenoid [WV82]; palatines small, narrow & tapered posteriorly [WV82]; palatines with broad anterior process articulating with proethmoid [WV82]; parasphenoid of moderate width [WV82]; parasphenoid with ventrolateral alar processes anteriorly, reaching mesopterygoids [WV82]; parasphenoid also with 1-2 pairs of more posterior alae, with carotid foramen variably located internal or ventral to more dorsal pair [WV82]; jaws short; mandible protrudes slightly beyond tip of upper jaw; deepest point of jaw at posterior end of dentary [WV82]; coronomeckelian of moderate to large size [WV82]; dorsal margin of jaw descend rapidly from posterior of dentary to quadrate articulation [WV82]; premaxilla with 3 rows of teeth, all ankylosed to jaw [WV82]; vomer with
6-9 teeth [WV82]; palatine with 2 tooth rows with 13 (lateral) & 6 (medial) teeth [WV82]; dentary with two rows of teeth, 22-25 in outer row & 9-14 in inner row [WV82]; epihyal triangular [WV82]; ceratohyal with prominent anterolateral processes [WV82]; hypohyals of roughly equal size [WV82]; urohyal broad, with well-developed dorsal keel [WV82]; branchiostegals spathiform, 3-5, with 1-2 attached to epihyal [WV82]; 4th epibranchial without uncinate process, vertically expanded distally [WV82] [R74]; but 4th external levator brachialis?) muscle present [JP97]; 4th epibranchial without levator process [JP97]; relict 5th, unossified epibranchial present [R74] [JP97] (contra [WV82]: not found); toothplates only on basibranchial 4 [WV82]; gill rakers longer than broad [WV82]; body robust, thick, little compressed; vertebrae 34-38 ([N94]: 32-37), with 18-21 precaudals [WV82]; centra longer than wide & constricted anteriorly (except last few) [WV82]; (pleural?) ribs articulate with centra & autogenous parapophyses absent [WV82]; neural and hemal arches fused to centra except last 2 [WV82]; no expanded neural spines, and supraneurals extend from 1st vertebra to 14th or 15th [WV82]; epineurals & epihemals present [WV82]; dorsal fin origin far anterior to anal fin [PB91]; 13-17 dorsal fin rays [PB91]; anal fin base half as long as dorsal [PB91]; 7-10 anal fin rays [PB91] [N94]; caudal fin rounded [PB91]; caudal fin with 18-20 rays; 5 hypurals, with 4+5 sometimes fused [WV82] (contra [R74]: 3 hypurals); 2 epurals [WV82]; 4-6 procurrent rays dorsally & ventrally, with 11-13 principal caudals [WV82],
8-9+ branched [R74]; extrascapular absent [JP97]; cleithrum & supracleithrum expanded [WV82]; urodermals reach anteriorly beyond first ural centrum [WV82]; large separate coracoid and 1 postcleithrum present [WV82]; 2-4 radials & 12(11 [PB91] [N94])-16 pectoral fin rays [WV82]; pelvic inserted just before origin of dorsal; pelvic girdle with elongate anterior pubic process & rounded illiac posterior plates, 6 (or 5 [N94])-7 fin rays [WV82] [PB91]; 30-37 large, cycloid lateral scales [PB91]; lateral line absent [PB91]; eats mostly insect larvae, copepods, "worms" [CS84]; preference for benthic foods [CS84]; unusual ability, especially in females, to hunt and metabolize food at very low temperatures [C+84]; air breathing possible through adaptations of swim bladder [C74].
posterior premaxillae not protractile; premaxilla without dorsal articular process [WV82]; maxilla deepest posteriorly [WV82]; supramaxilla absent [WV82]; frontals terminate anteriorly below posterior 1/3rd of proethmoid [WV82]; frontal comes to a point over sphenotics [WV82]; frontal transverse ridge absent [WV82]; frontals overlie variable?) portion of supraoccipital [WV82]; parietals small (intermediate between Novumbra and Dallia) [WV82]; posttemporal small [WV82]; suspensorium short & deep [WV82]; symplectic not straight [W84]; preopercular with ventral arm larger than dorsal [WV82]; opercular with small lateral process, stout reinforcing rays, and slight posterior excavation [WV82]; subopercular & interopercular both triangular, of equal size and lack processes [WV82]; basioccipital with anteriorly-directed median spur (unique to Umbra) [WV82]; exoccipitals extended in posterodorsal- anteroventral axis [WV82]; sphenotic with lateral process at apex for origin of m. levator arcus palatini [WV82]; alisphenoids do not meet on the midline [WV82]; alisphenoid with large, ventrally- directed flange [WV82]; proethmoid anterior margin partially endochondral, curved ventrolaterally [WV82]; proethmoid anterior margin expanded & articulates with dorsolateral surface of premaxilla [WV82]; proethmoid thickened & grooved posteriorly to cap anterior end of rostral cartilage & articulate via cartilaginous joint with anteromedial margin of palatines [WV82]; vomer small, generally antero-posteriorly oriented & located variably ventral or under parasphenoid [WV82]; palatines small, narrow & tapered posteriorly [WV82]; palatines with broad anterior process articulating with proethmoid [WV82]; parasphenoid of moderate width [WV82]; parasphenoid with ventrolateral alar processes anteriorly, reaching mesopterygoids [WV82]; parasphenoid also with 1-2 pairs of more posterior alae, with carotid foramen variably located internal or ventral to more dorsal pair [WV82]; jaws short; mandible protrudes slightly beyond tip of upper jaw; deepest point of jaw at posterior end of dentary [WV82]; coronomeckelian of moderate to large size [WV82]; dorsal margin of jaw descend rapidly from posterior of dentary to quadrate articulation [WV82]; premaxilla with 3 rows of teeth, all ankylosed to jaw [WV82]; vomer with
6-9 teeth [WV82]; palatine with 2 tooth rows with 13 (lateral) & 6 (medial) teeth [WV82]; dentary with two rows of teeth, 22-25 in outer row & 9-14 in inner row [WV82]; epihyal triangular [WV82]; ceratohyal with prominent anterolateral processes [WV82]; hypohyals of roughly equal size [WV82]; urohyal broad, with well-developed dorsal keel [WV82]; branchiostegals spathiform, 3-5, with 1-2 attached to epihyal [WV82]; 4th epibranchial without uncinate process, vertically expanded distally [WV82] [R74]; but 4th external levator brachialis?) muscle present [JP97]; 4th epibranchial without levator process [JP97]; relict 5th, unossified epibranchial present [R74] [JP97] (contra [WV82]: not found); toothplates only on basibranchial 4 [WV82]; gill rakers longer than broad [WV82]; body robust, thick, little compressed; vertebrae 34-38 ([N94]: 32-37), with 18-21 precaudals [WV82]; centra longer than wide & constricted anteriorly (except last few) [WV82]; (pleural?) ribs articulate with centra & autogenous parapophyses absent [WV82]; neural and hemal arches fused to centra except last 2 [WV82]; no expanded neural spines, and supraneurals extend from 1st vertebra to 14th or 15th [WV82]; epineurals & epihemals present [WV82]; dorsal fin origin far anterior to anal fin [PB91]; 13-17 dorsal fin rays [PB91]; anal fin base half as long as dorsal [PB91]; 7-10 anal fin rays [PB91] [N94]; caudal fin rounded [PB91]; caudal fin with 18-20 rays; 5 hypurals, with 4+5 sometimes fused [WV82] (contra [R74]: 3 hypurals); 2 epurals [WV82]; 4-6 procurrent rays dorsally & ventrally, with 11-13 principal caudals [WV82],
8-9+ branched [R74]; extrascapular absent [JP97]; cleithrum & supracleithrum expanded [WV82]; urodermals reach anteriorly beyond first ural centrum [WV82]; large separate coracoid and 1 postcleithrum present [WV82]; 2-4 radials & 12(11 [PB91] [N94])-16 pectoral fin rays [WV82]; pelvic inserted just before origin of dorsal; pelvic girdle with elongate anterior pubic process & rounded illiac posterior plates, 6 (or 5 [N94])-7 fin rays [WV82] [PB91]; 30-37 large, cycloid lateral scales [PB91]; lateral line absent [PB91]; eats mostly insect larvae, copepods, "worms" [CS84]; preference for benthic foods [CS84]; unusual ability, especially in females, to hunt and metabolize food at very low temperatures [C+84]; air breathing possible through adaptations of swim bladder [C74].
Notes: Some of the meristic characters are from [PB91] and have been widely republished elsewhere. However, Page & Burr were not concerned with the European species, E. krameri, which may extend some of these ranges.
Links: Mudminnow Family, Umbridae, central mudminnow, umbra limi; Master Index of Freshwater Fishes (U - V); Umbra krameri; Central Mudminnow; Umbra limi; Nonindigenous fishes - Umbra limi; Central Mudminnow; More Central Mudminnow Info; Central Mudminnow; NatureServe Explorer Species Index- Genus UMBRA.
References: Chilton et al. (1984) [C+84]; Colgan & Silburt (1984) [CS84]; Crawford 1974) [C74]; Johnson & Patterson (1997) [JP97]; Page & Burr 1991) [PB91]; Rosen (1974) [R74]; Wilson (1984) [W84]; Wilson & Veilleux (1982) [WV82]. ATW030412.
 Dallia: D. pectoralis Bean 1880 (not 1879) (Alaskan blackfish). D. admirabilis Chereshnev 1980 and D. delictissima Smitt 1881 are possibly junior synonyms. Genus is coextensive with Dallidae, Dallinae, and Xenomi.
Dallia: D. pectoralis Bean 1880 (not 1879) (Alaskan blackfish). D. admirabilis Chereshnev 1980 and D. delictissima Smitt 1881 are possibly junior synonyms. Genus is coextensive with Dallidae, Dallinae, and Xenomi.
Range: from the Miocene [K+86] of Alaska, Bering Sea islands & Siberia.
Phylogeny: Esociformes :: Novumbra + Esocidae) + *.
Characters: to 33 cm [PB91]; snout short, flattened [PB91]; premaxilla borders >50% of gape [WV82]; premaxilla with prominent articular process on dorsal margin fitting against head of maxilla [WV82]; maxilla slender, tapering posteriorly, with single supramaxilla [WV82]; frontal ![Dallia paired fin girdles from [WV82]](images/DalliaFinGirdles.gif) extends anteriorly only to lateral ethmoids [WV82]; frontals separated for several centimeters anteriorly [WV82]; frontals marked dorsally by ridges [WV82]; frontal comes to bluntly rounded end over sphenotics [WV82]; frontal transverse ridge present, with shallow posttemporal fossa behind ridge [WV82]; frontal overlies only a small portion of the supraoccipital [WV82]; parietals largest of basal esociforms, covering lateral quarter of supraoccipital [WV82]; supraoccipital with posterior crest somewhat reduced [WV82]; supraoccipital with posterior notch on ventral surface [WV82]; posttemporal slender [WV82]; suspensorium long & shallow [WV82]; symplectic nearly straight [W84]; preopercular expanded at junction of arms, with dorsal arm short [WV82]; opercular small, subtriangular, & notched posteriorly [WV82]; subopercular as in Novumbra, but notched anteriorly for interopercular [WV82]; interopercular narrow anteriorly & forked posteriorly [WV82]; exoccipitals antero-posteriorly long [WV82]; sphenotic with lateral process at apex for origin of m. levator arcus palatini [WV82]; alisphenoids meet on the midline [WV82]; proethmoid triangular [WV82]; proethmoid anterior margin partially endochondral, curved ventrolaterally [WV82]; proethmoid anterior margin expanded & articulates with dorsolateral surface of premaxilla [WV82]; proethmoid thickened & grooved posteriorly to cap anterior end of rostral cartilage & articulate via cartilaginous joint with anteromedial margin of palatines [WV82]; palatines long, with dorsal groove flanked by medial & dorsal flanges [WV82]; palatine with anterior articular facet for proethmoid joint [WV82]; palatine tapers posteriorly to point near anterodorsal corner of quadrate [WV82]; mesopterygoid rounded anteriorly [WV82]; parasphenoid very broad, not waisted, and lacking alae or carotid foramina [WV82]; parasphenoid, anterior portion concave ventrally [WV82]; vomer transversely elongated [WV82]; mandible long & gracile, with deepest point at anterior end of anguloarticular [WV82]; coronomeckelian moderate size & located midway along anguloarticular [WV82]; premaxilla with single row of 24-28 teeth [WV82]; vomer with 11-13 teeth located anteriorly [WV82]; palatine with single row of 14-15 teeth [WV82]; dentary with single row of ~17 teeth [WV82]; hyomandibular shaft short & opercular process long [WV82]; epihyal rounded posteriorly [WV82]; urohyal small & serrated posteriorly [WV82]; branchiostegals 7-9, acinaciform, with 3 attached to epihyal [WV82]; 4th epibranchial with (reduced) uncinate process [WV82] [R74]; but 4th external levator (brachialis?) muscle present [JP97]; 4th epibranchial without levator process [JP97]; 5th epibranchial absent [JP97]; branchial toothplates absent on basihyal, but present on basibranchials 3-4 [WV82]; gill
extends anteriorly only to lateral ethmoids [WV82]; frontals separated for several centimeters anteriorly [WV82]; frontals marked dorsally by ridges [WV82]; frontal comes to bluntly rounded end over sphenotics [WV82]; frontal transverse ridge present, with shallow posttemporal fossa behind ridge [WV82]; frontal overlies only a small portion of the supraoccipital [WV82]; parietals largest of basal esociforms, covering lateral quarter of supraoccipital [WV82]; supraoccipital with posterior crest somewhat reduced [WV82]; supraoccipital with posterior notch on ventral surface [WV82]; posttemporal slender [WV82]; suspensorium long & shallow [WV82]; symplectic nearly straight [W84]; preopercular expanded at junction of arms, with dorsal arm short [WV82]; opercular small, subtriangular, & notched posteriorly [WV82]; subopercular as in Novumbra, but notched anteriorly for interopercular [WV82]; interopercular narrow anteriorly & forked posteriorly [WV82]; exoccipitals antero-posteriorly long [WV82]; sphenotic with lateral process at apex for origin of m. levator arcus palatini [WV82]; alisphenoids meet on the midline [WV82]; proethmoid triangular [WV82]; proethmoid anterior margin partially endochondral, curved ventrolaterally [WV82]; proethmoid anterior margin expanded & articulates with dorsolateral surface of premaxilla [WV82]; proethmoid thickened & grooved posteriorly to cap anterior end of rostral cartilage & articulate via cartilaginous joint with anteromedial margin of palatines [WV82]; palatines long, with dorsal groove flanked by medial & dorsal flanges [WV82]; palatine with anterior articular facet for proethmoid joint [WV82]; palatine tapers posteriorly to point near anterodorsal corner of quadrate [WV82]; mesopterygoid rounded anteriorly [WV82]; parasphenoid very broad, not waisted, and lacking alae or carotid foramina [WV82]; parasphenoid, anterior portion concave ventrally [WV82]; vomer transversely elongated [WV82]; mandible long & gracile, with deepest point at anterior end of anguloarticular [WV82]; coronomeckelian moderate size & located midway along anguloarticular [WV82]; premaxilla with single row of 24-28 teeth [WV82]; vomer with 11-13 teeth located anteriorly [WV82]; palatine with single row of 14-15 teeth [WV82]; dentary with single row of ~17 teeth [WV82]; hyomandibular shaft short & opercular process long [WV82]; epihyal rounded posteriorly [WV82]; urohyal small & serrated posteriorly [WV82]; branchiostegals 7-9, acinaciform, with 3 attached to epihyal [WV82]; 4th epibranchial with (reduced) uncinate process [WV82] [R74]; but 4th external levator (brachialis?) muscle present [JP97]; 4th epibranchial without levator process [JP97]; 5th epibranchial absent [JP97]; branchial toothplates absent on basihyal, but present on basibranchials 3-4 [WV82]; gill ![Dallia caudal skeleton [WV82]](images/DalliaCaudalSkel.gif) rakers stout, triangular, & finely toothed [WV82]; vertebrae 40-41 (or 42 [N94]]) with 19-20 precaudals [WV82]; neural & hemal arches fused to centra except posterior 2-3 [WV82]; anterior neural spines present, but not fused at midline [WV82]; precaudals with pleural ribs attached to separate parapophyseal ossifications [WV82]; epineurals absent [WV82]; anterior 3 neural spines expanded distally, followed by 19-21 spines bearing sigmoid supraneurals [WV82]; dorsal and anal fins located posteriorly; anal more or less exactly under dorsal, with equal fin base length [PB91]; fin spines absent; dorsal soft rays 10-14 [PB91] [N94]; anal soft rays: 11-16 [PB91] ([N94]: 12-16); caudal broad and rounded [PB91]; caudal fin large, with highly variable skeleton [WV82]; preurals 1-2 may have single neural arch [R74]; 3-5 hypurals with last 2 frequently fused (contra [R74]: 1-3 hypurals, counting fusions as 1) and 1st few of same size [WV82]; accessory hemal spine present [WV82]; 0-1 epurals [WV82] (contra [R74]: 1 epural); 6-8 procurrent rays dorsal & ventral, with 20-22 principals caudals (18-20 branched) [WV82]; urodermals don't reach anteriorly beyond first ural centrum [R74] [WV82]; extrascapular absent [JP97]; edge of pectorals rounded; separate coracoid absent & postcleithrum absent [WV82]; ossified Baudelot's ligament present [WV82] [N94]; broad, unossified radial plate with 30-36 rays in the pectorals [WV82] (compare [PB91] & [N94]: 32-36 rays); pectoral fins broadly rounded [PB91]; pelvics very small and located just before anus; pelvic girdle with small rodlike pubic [WV82]; pelvic fins tiny, with 2-3 fin rays [PB91] [N94]: rare individuals with 0-1); lateral line with minute pores [PB91]; 76 to 100 tiny scales in midlateral series [PB91]; usually found in heavily vegetated swamps and ponds [PB91]; occasionally in medium to large rivers and lakes with abundant vegetation [PB91]; migrations appear to be limited to inshore or upstream movements to spawning grounds in the spring and (presumably) reverse migrations to deeper water in the fall; tolerates extreme cold, including some freezing; uses esophagus as auxiliary breathing organ [C74].
rakers stout, triangular, & finely toothed [WV82]; vertebrae 40-41 (or 42 [N94]]) with 19-20 precaudals [WV82]; neural & hemal arches fused to centra except posterior 2-3 [WV82]; anterior neural spines present, but not fused at midline [WV82]; precaudals with pleural ribs attached to separate parapophyseal ossifications [WV82]; epineurals absent [WV82]; anterior 3 neural spines expanded distally, followed by 19-21 spines bearing sigmoid supraneurals [WV82]; dorsal and anal fins located posteriorly; anal more or less exactly under dorsal, with equal fin base length [PB91]; fin spines absent; dorsal soft rays 10-14 [PB91] [N94]; anal soft rays: 11-16 [PB91] ([N94]: 12-16); caudal broad and rounded [PB91]; caudal fin large, with highly variable skeleton [WV82]; preurals 1-2 may have single neural arch [R74]; 3-5 hypurals with last 2 frequently fused (contra [R74]: 1-3 hypurals, counting fusions as 1) and 1st few of same size [WV82]; accessory hemal spine present [WV82]; 0-1 epurals [WV82] (contra [R74]: 1 epural); 6-8 procurrent rays dorsal & ventral, with 20-22 principals caudals (18-20 branched) [WV82]; urodermals don't reach anteriorly beyond first ural centrum [R74] [WV82]; extrascapular absent [JP97]; edge of pectorals rounded; separate coracoid absent & postcleithrum absent [WV82]; ossified Baudelot's ligament present [WV82] [N94]; broad, unossified radial plate with 30-36 rays in the pectorals [WV82] (compare [PB91] & [N94]: 32-36 rays); pectoral fins broadly rounded [PB91]; pelvics very small and located just before anus; pelvic girdle with small rodlike pubic [WV82]; pelvic fins tiny, with 2-3 fin rays [PB91] [N94]: rare individuals with 0-1); lateral line with minute pores [PB91]; 76 to 100 tiny scales in midlateral series [PB91]; usually found in heavily vegetated swamps and ponds [PB91]; occasionally in medium to large rivers and lakes with abundant vegetation [PB91]; migrations appear to be limited to inshore or upstream movements to spawning grounds in the spring and (presumably) reverse migrations to deeper water in the fall; tolerates extreme cold, including some freezing; uses esophagus as auxiliary breathing organ [C74].
Links: Common Name - Search Result, SPECIES; NatureServe Explorer Species Index- Genus DALLIA; NAS - Species FactSheet.
References: Crawford 1974) [C74]; Johnson & Patterson (1997) [JP97]; Page & Burr 1991) [PB91]; Kettler et al. 1986) [K+86]; Rosen (1974) [R74]; Wilson (1984) [W84]; Wilson & Veilleux (1982) [WV82]. ATW030412.