 The Whale's Tale: Darwinian Destiny or Fluke of Chance?
The Whale's Tale: Darwinian Destiny or Fluke of Chance?| Cetartiodactyla | ||
| The Vertebrates | Basilosauridae |
| Vertebrates Home | Vertebrate | Vertebrate |
|
Abbreviated Dendrogram
Laurasiatheria
├─Ferae
└─┬─Perissodactyla
│
└─Cetartiodactyla
├─Cetacea
│ ├─Pakicetidae
│ └─Protocetidae
│ ├─Georgiacetus
│ └─Basilosauridae
│ ├─Basilosaurus
│ └─┬─Dorudon
│ └─Autoceta
└─Artiodactyla
├─Suina
│ ├─Hippopotamidae
│ └─Suoidea
└─Selenodontia
├─Camelidae
└─Ruminantia
├─Giraffoidea
└─┬─Bovoidea
└─Cervoidea
|
Contents
Overview |
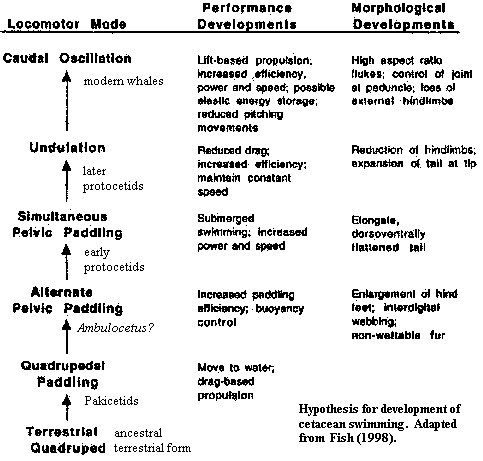
The Whale's Tale: Darwinian Destiny or Fluke of Chance?Here we attempt to tie up several themes we have developed during this somewhat extended treatment of Eocene whales. The problem we will address is relates to whale locomotion at the end of the Eocene. We understand quite well how whales swam at the beginning of the Eocene. There are many terrestrial mammals who spend considerable time in water, and their swimming styles are reasonably well known. Many whales are alive today, and they swim by dorsoventral undulation of their tail flukes. The problem comes in changing from the limb-powered locomotion of the earliest whales and basal protocetids to the fluke-propelled locomotion of modern whales. The current best guess in this area was developed by Fish 1998) [1]. We will argue that Fish's theory is slightly incomplete and that the morphotype of Basilosaurus is not an aberration or sideline, as argued by Buchholtz 1998), but a necessary intermediate form.
This issue has broader implications for our assertion that, at least until Late Oligocene times, whales were evolving in a Darwinian fashion. We have asserted that one can treat all Eocene Cetacea essentially as a single population moving through phylospace along a smooth morphocline, propelled by a reasonably constant set of selective forces. Basilosaurus and its locomotor mode are an important test of this hypothesis. If Basilosaurus is a uniquely derived evolutionary dead end, we are probably wrong. If it was a mechanically necessary intermediate form, then we are vindicated once more [2].
Fish's hypothesis is shown on the right. In essence, the Cetacea progress from dog-padding to hindlimb-powered swimming involving first alternate, and then simultaneous use of the legs. He derived this idea from a critical theoretical insight: locomotor activity is inherently very conservative. Paddling motions are instinctive because they are very close to normal terrestrial quadrupedal motion. Simultaneous pelvic paddling is very close to the bounding run of terrestrial hunters. The latter is primarily an underwater swimming mode. As the organism spends more time under water, it also becomes advantageous to develop a wide tail which undulates up and down. Eventually, the tail powered component becomes the more important, and the hind limbs are reduced and eliminated because they contribute to drag.
We will return to this hypothetical sequence in a moment. However, we pause momentarily to review in a bit more detail,  the method by which flukes operate. As Fish discusses, flukes are essentially airfoils, like the wing of a plane. In cross-section, they have precisely this tear-drop shape. Now: the function of an airfoil wing on a plane is not to provide motive power. It is to (a) maintain lift and (b) minimize turbulence, which creates drag. These are slightly different functions. The second function is best served by a completely symmetrical airfoil -- completely tear-drop shaped -- which faces directly into the line of motion. The medium, whether air or water, flows smoothly over top and bottom, and the flow lines converge toward their initial spacing and position.
the method by which flukes operate. As Fish discusses, flukes are essentially airfoils, like the wing of a plane. In cross-section, they have precisely this tear-drop shape. Now: the function of an airfoil wing on a plane is not to provide motive power. It is to (a) maintain lift and (b) minimize turbulence, which creates drag. These are slightly different functions. The second function is best served by a completely symmetrical airfoil -- completely tear-drop shaped -- which faces directly into the line of motion. The medium, whether air or water, flows smoothly over top and bottom, and the flow lines converge toward their initial spacing and position.
The spacing between the flow lines relates to the density of the medium. Since the flow is symmetrical in the minimum drag case, the pressure is the same on both sides of the foil, and there is no net lift. However in the second case, if we tilt the airfoil or make it asymmetrical, the result is quite different. The flow lines are compressed, and the pressure beneath the foil is increased. The flow lines above the foil are separated, and so pressure above the foil decreased. In short, there is a significant pressure difference above and below the foil and therefore a net positive lift. However, the surface area of the foil directly opposed to the line of motion increases, which creates drag. Worse, as the streams bend to equalize pressure behind the foil, and may set up a turbulent gyre further slowing the foil by induced drag.
The optimum angle of attack (most lift for least drag) depends on the density of the medium and the forward velocity. The computation is further complicated for a dorsoventrally moving foil, like a dolphin's tail. We don't need to get into the math. It is only necessary to observe that the best angle of attack is a critical and constantly changing parameter. Dolphins and modern whales are able to manage the angle of attack by means of a "ball vertebra" in the caudal peduncle (the constricted portion of the tail just anterior to the fluke) which allows them to trim the angle of the fluke separately from the angle of the tail.
 Now, while still keeping all this physics in your head, consider the problem of evolving from a bounding sort of action, using both legs at once, to caudal undulation using a fluke. Here we are, smoothly bringing the tail down with the fluke adjusted to just the right angle of attack, when -- bang! -- suddenly our legs kick back with a burst of acceleration. Two things go wrong. First, the angle of attack, which was right for our initial velocity, is wrong for the velocity post-kick. Second, an undulating tail is, by definition, usually off the centerline of the body. It will therefore act as a rudder in response to a quick acceleration. The result is that we'll go tearing off at a new angle, for which our flukes are not correctly trimmed. So Fish's smooth step from simultaneous pelvic paddling to caudal undulation masks an important discontinuity [3].
Now, while still keeping all this physics in your head, consider the problem of evolving from a bounding sort of action, using both legs at once, to caudal undulation using a fluke. Here we are, smoothly bringing the tail down with the fluke adjusted to just the right angle of attack, when -- bang! -- suddenly our legs kick back with a burst of acceleration. Two things go wrong. First, the angle of attack, which was right for our initial velocity, is wrong for the velocity post-kick. Second, an undulating tail is, by definition, usually off the centerline of the body. It will therefore act as a rudder in response to a quick acceleration. The result is that we'll go tearing off at a new angle, for which our flukes are not correctly trimmed. So Fish's smooth step from simultaneous pelvic paddling to caudal undulation masks an important discontinuity [3].
Fish's own, extensive work with modern analogues of early whales provides some insights on this dilemma. One of the most aquatic of the otter family is Pteronura, the giant river otter of South America. Fish repeatedly refers to Pteronura because it uses both simultaneous hindlimb propulsion and undulation of its long, flat tail. For his purposes, the integration of these two modes is immaterial. However, for us, it is critical. As it turns out, Pteronura does not even attempt to integrate the two swimming modes. It uses its webbed feet for slow maneuvering. It uses its tail for fast travel. It does not use both at once.
 Now, with these apparently random bits of information, we are at last properly armed to resist Prof. Buchholtz's assault on Basilosaurus. Buchholtz is primarily focused on the axial skeleton. She envisions the Eocene evolution of whales as involving the following steps:
Now, with these apparently random bits of information, we are at last properly armed to resist Prof. Buchholtz's assault on Basilosaurus. Buchholtz is primarily focused on the axial skeleton. She envisions the Eocene evolution of whales as involving the following steps:
1) shortening and stiffening the neck and anterior spine to create a stiff, non-undulating, cervico-thoracic region.
2) progressive elongation of the lumbar vertebrae, with incorporation of the former sacrals (no longer attached to the pelvis), anterior caudals and posterior thoracics into an undulating torso. This is accompanied by a strong tendency for the body to become more sleek.
3) The propulsive force is initially provided by undulation of the flattened torso itself, with a gradual posterior shift of the region undergoing maximum displacement, ultimately culminating in tail-based propulsion using a fluke.
To Buchholtz, Basilosaurus is simply too long. The vertebral centra are all very long and do not increase much in length along the torso. The tail is also long. Although Basilosaurus possessed a fluke and peduncle, Buchholtz dismisses these features because they are too short relative to body length. Buchholtz (1998: 346). Thus, she asserts, locomotion in Basilosaurus was probably anguilliform, and the genus was an evolutionary sideline with no living counterparts.
These are powerful arguments. But we "are constant as the Northern Star, of whose true fixed and resting quality there is no fellow in the firmament." Why do all known protocetids seem to be headed in the direction of longer, leaner body forms throughout the Eocene? If Basilosaurus is a mammalian eel, why does it have a well-developed stiffened thoracic region? Why would it have a fluke at all? Why would an anguilliform swimmer have those big forelimbs?
 Let's start over again from scratch. Fish shows that early protocetids would naturally go to a two-legged propulsion style because it is similar to a bounding run. What else do we know about bounding mammalian runners? They, too, are typically long and lean. The back legs are brought well forward with the spine curved. The hind legs kick back, and the body extends hence the advantage of length), with the forelimbs extended. The fore limbs dig in and help momentum haul the back end into the curled, poised starting position for the next spring. So what are the protocetid's forelimbs doing underwater? Perhaps much the same thing. If so, the motion would look remarkably like the paddling motion of Stellar's Sea Lion, Eumetopias, whose forelimb swimming motions were discussed in connection with the Plesiosauria. Buchholtz places much emphasis on vertebral charts which track centra dimensions along the spine. It is interesting, and possibly relevant, that the chart for Eumetopias looks much like the chart for Basilosaurus, but for the greater elongation of the centra in the whale.
Let's start over again from scratch. Fish shows that early protocetids would naturally go to a two-legged propulsion style because it is similar to a bounding run. What else do we know about bounding mammalian runners? They, too, are typically long and lean. The back legs are brought well forward with the spine curved. The hind legs kick back, and the body extends hence the advantage of length), with the forelimbs extended. The fore limbs dig in and help momentum haul the back end into the curled, poised starting position for the next spring. So what are the protocetid's forelimbs doing underwater? Perhaps much the same thing. If so, the motion would look remarkably like the paddling motion of Stellar's Sea Lion, Eumetopias, whose forelimb swimming motions were discussed in connection with the Plesiosauria. Buchholtz places much emphasis on vertebral charts which track centra dimensions along the spine. It is interesting, and possibly relevant, that the chart for Eumetopias looks much like the chart for Basilosaurus, but for the greater elongation of the centra in the whale.
As for the length of centra, this is undoubtedly (as Buchholtz observes) related to greater flexibility in the spine. But there are other purposes for flexibility than eel-like undulation. For example, turning at speed in water is a difficult proposition. A long-bodied form (like a pike, for example) is able to turn quickly using its body as an enormous rudder. Far from indicating slow movement, the length of Basilosaurus may be an indicator of high-speed pursuit behavior. This would be far more consistent with what we know about protocetid diets and their place in the food chain (see the Georgiacetus essay).
It would be nice, at this point, to reach into our magic hat and -- presto! -- pull out a complete theory of locomotion in Basilosaurus. We are unable to oblige. However, we do have enough information to conclude that Basilosaurus cannot yet be consigned to the trash heap of hopeless maladaptations and evolutionary dead ends. We can sum up as follows:
1) The thoracic region was stout and stiff. The fore limbs could, and probably did, participate in at least some locomotor modes in addition to providing guidance.
2) The body may originally have been lengthened to make best "bounding" use of smaller legs, creating a smoother hydrodynamic outline for coasting between "leaps." Whatever the locomotor mode, a long body would be advantageous for quick turns and ambush predation.
2) Anguilliform motion may have been possible. There is no reason to assume it was exclusive, or that the lengthening of lumbar centra -- observed in all later protocetids -- was solely, or even primarily related to eel-like motion.
3) Caudal undulation probably began independently of two-legged locomotion. It was an alternate mode, rather than an extension of what early protocetids were already doing. Basilosaurus was probably capable of tail-based propulsion, using a relatively long tail with only a small fluke. Perhaps, as in long-bodied fishes, caudal undulation was used as a low-efficiency, but very high-speed, locomotor mode for ambush predation.
In short, Basilosaurus may be no more unique or specialized than other late protocetids. It represents a grade between Georgiacetus and Dorudon in which the hind limb was almost completely eliminated as an organ of propulsion. If the animal seems a bit awkward, this is only because it was an awkward stage in transition between two incompatible locomotor modes. As caudal undulation, particularly the fluke, developed further, it would be possible to shorten the tail and body and eliminate the role of the forelimbs in locomotion. ATW030202.
[1] Since Prof. Frank Fish has undoubtedly heard every possible joke related to his name in connection with his work on whales, we will leave all such remarks as an exercise for the reader. [2] Obviously we would not be "proved correct." Hypotheses cannot be proven. They can only be tested and disproven. Frankly, we are personally unhappy with the idea of Eocene whales paddling around out there, raising hell with the techniques of Hennig and the theories of Gould. However, this seems to be the case. [3] We've given the matter no serious thought, but might this also be a problem for ground-up theories of dinosaur flight?
 Basilosauridae: Basiloterus
Basilosauridae: Basiloterus
Range: from the Middle Eocene (Bartonian) [G98] [U98]
Protocetidae : Georgiacetus + * : Basilosaurus + (Dorudon + Autoceta)
nostrils not terminal [Z98]; nares open above P1 or further back [T98]; primitively, no telescoping of either occipital or rostral elements over braincase [U98]; $? pterygoid sinuses [FB94] [GU98] [KF97]; "peribullar air sinuses" [U98]; tubular portion of ectotympanic contribution to external auditory meatus absent [L98]; partially isolated periotics [GU98]; dense tympanic bullae [GU98]; conical apophysis of bulla fully developed [L98$]; bulla ectotympanic annulus vestigial or absent [L98$]; anterior process (processus tubarius) of bulla articulates extensively with petrosal [L98$]; large mandibular canals with lateral acoustic fenestrae [GU98]; $? multiple accessory denticles on molars & premolars [U98] [KF97]; protocone reduced [T98]; $? loss of M3 [U98]; short cervical vertebrae [T+01] [U98]; post-cervical vertebrae tend to be of similar length, except distal caudals [U98]; number of lumbar vertebrae increased to about 20 [U98]; unfused sacral vertebrae [T+01] [U98]; no contact between pelvis & vertebrae [T+01] [U98]; short tail with ball & socket vertebral articulations [T+01] [1]; mid-caudals shorten abruptly, suggesting tail fluke [U98]; scapula broad & fan-shaped [U98]; scapula with anterior acromion process and small supraspinous fossa [T+01] [U98]; coracoid process oriented anteriorly [U98]; humerus increasingly robust [U98]; humerus much longer than radius [G98] (contra [U98] humerus same length as radius & ulna); humerus with large hemispherical head & well-developed proximal muscle attachments [U98]; humerus with strong deltopectoral crest [U98]; single humeral trochlea for ulna & radius [U98]; ulna with large and transversely flat olecranon [T+01] [U98]; ulnar shaft also flat along entire length [U98]; radius head roughly circular and only slightly concave [U98]; radius articulates with ulna along flat surface [U98]; radius shaft flattened distally [U98]; carpals flattened in plane of flipper [U98]; carpal articular surfaces do not extend onto dorsal or palmar surfaces of carpals [U98]; trapezoid & magnum fused [U98]; forelimbs paddle-shaped [T+01]; forelimbs relatively narrow [Z98]; manus pentadactyl & without hyperphalangy [U98]; pelvis not attached to spine [G98]; pelvis very short [U98]; hind limbs vestigial and internal [T+01].
 Note:
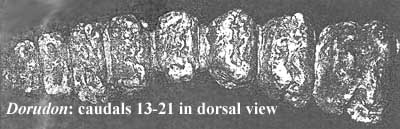
[1] Image from [U98]. Yes, we also only count 8 vertebrae. That's just the way it was labeled in the original. The abrupt change in shape and thickness noted by Uhen is quite clear, and this indeed suggests the presence of some novel caudal structure. However, the "ball vertebra" described by [T+01] as a synapomorphy of Basilosauridae is simply not present, nor is there a caudal peduncle. So, this is not a fluke such as a whale might have today. In fact, it might not make any mechanical sense for an elongate swimmer like Basilosaurus to have a fluke if it were actually a anguilliform swimmer (but see discussion above). On the physical evidence, the caudal structure could, as easily, represent a beaver-like tail for nursery building, for communication, or for herding fish. Of course, there's no evidence for any of that, either. But why assume a structure that makes no mechanical sense and is not supported by the fossil record, simply because an organism's distant cousins had it 30 My later? Just because Homo habilis had strong legs and long arms doesn't mean that it rode a bicycle.
Note:
[1] Image from [U98]. Yes, we also only count 8 vertebrae. That's just the way it was labeled in the original. The abrupt change in shape and thickness noted by Uhen is quite clear, and this indeed suggests the presence of some novel caudal structure. However, the "ball vertebra" described by [T+01] as a synapomorphy of Basilosauridae is simply not present, nor is there a caudal peduncle. So, this is not a fluke such as a whale might have today. In fact, it might not make any mechanical sense for an elongate swimmer like Basilosaurus to have a fluke if it were actually a anguilliform swimmer (but see discussion above). On the physical evidence, the caudal structure could, as easily, represent a beaver-like tail for nursery building, for communication, or for herding fish. Of course, there's no evidence for any of that, either. But why assume a structure that makes no mechanical sense and is not supported by the fossil record, simply because an organism's distant cousins had it 30 My later? Just because Homo habilis had strong legs and long arms doesn't mean that it rode a bicycle.
Links: Whale Transitional Fossil Evidence; Digital \I\ Designs - Cenozoic Images - Basilosaurus Dance; Cetaceans.
References: Fordyce & Barnes (1994) [FB94]; Gingerich 1998) [G98]; Gingerich & Uhen (1998) [GU98]; Köhler & Fordyce (1997) [KF87]; Luo (1998) [L98]; Thewissen 1998) [T98]; Thewissen et al. (2001) [T+01]; Uhen 1998) [U98]; Zimmer 1998) [Z98]. ATW030125.
 Basilosaurus: Harlan 1834. B. cetoides Harlan 1834; B. drazindai Gingerich et al. 1997; B. isis Andrews 1904.
Basilosaurus: Harlan 1834. B. cetoides Harlan 1834; B. drazindai Gingerich et al. 1997; B. isis Andrews 1904.
Range: Middle Eocene (Bartonian) to Late Eocene (Priabonian) [U98] South Asia, Africa, Europe & North America, possibly New Zealand [U98].
Phylogeny: Basilosauridae : (Dorudon + Autoceta) + *.
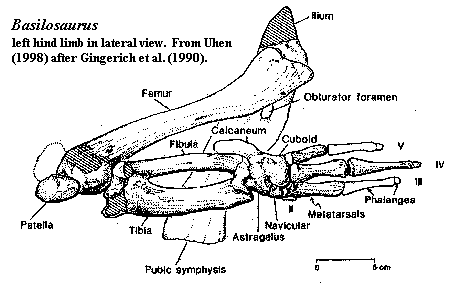 Characters: 5-25m; body long and thin [Z98] [B98]; nostrils almost terminal [Z98]; dense tympanic bullae [GU98]; pterygoid sinuses [FB94] [GU98] partially isolated periotics [GU98]; large mandibular canals with lateral acoustic fenestrae [GU98]; cheek teeth retain complex morphology and functional occlusion [FB94] [GU98]; heavy wear facets (food was chewed) [GU98]; molars and premolars with 2 roots [C88]; loss of M3; elongate, with light, cancelous vertebrae probably filled with oil [G98] [Z+00]; short cervical vertebrae [T+01]; vertebral centra generally long and barrel-shaped [Z98]; elongated post-cervical centra [G98] [U98]; thoracic and lumbar vertebrae approximately same length [T+01]; ~15 thoracics, with last 7 lumbarized [B98]; thoracics moderately wide (width/height 1.3-1.5) [B98]; thoracic ribs strongly pachyostosed [M98]; long torso all involved in undulation [B98]; all lumbarized vertebrae elongated, with length ~twice height and width only slightly greater than height [B98]; 15 lumbars (includes "sacrals") [B98]; unfused sacral vertebrae [T+01]; sacro-illiac "joint" absent [T+01]; short tail with ball & socket vertebral articulations [T+01]; anterior 6 caudals also lumbarized [B98]; probable peduncle & tail fluke based on midcaudal reduction in centrum length & centra wider than tall [B98]; broad, fan-shaped scapulae [T+01]; scapula with anterior acromion process and small supraspinous fossa [T+01]; humerus much longer than radius [G98]; elbow joint primitively mobile [G98]; ulna with large and transversely flat olecranon [T+01]; forelimbs paddle-shaped [T+01]; forelimbs relatively narrow [Z98]; simplified pelvis joined only ventrally [Z98]; pelvis shorter than any thoracic vertebra [U98]; pelvis not attached to spine [G98]; pubis greatly elongated [U98]; pubis located anterior to obturator foramen (implying that pelvis has been rotated counterclockwise, assuming a right lateral view) [U98]; hind limbs vestigial and internal [T+01]; hind limbs could assume only two positions, used as copulatory guide? [Z98]; femurs small & gracile, shorter than innominate [U98]; femur containing extremely dense bone in central shaft (diaphyseal bone) [M98]; proximal femur sometimes malformed [U98]; femur with patellar groove [U98] patella present, ovate & thick [U98]; tibia & fibula fused [U98]; tarsals variably fused [U98]; pes I absent, pes II vestigial [U98]; may have retained terrestrial habit seasonally; long, flat plateau in vertebral morphology indicates anguilliform motion probably first cosmopolitan (not just Tethys) cetacean group [Z98]; anguilliform swimmers, using lateral undulation, who lived primarily on the surface waters [G98] (contra [B98]: anguilliform swimmers with dorsoventral undulation).
Characters: 5-25m; body long and thin [Z98] [B98]; nostrils almost terminal [Z98]; dense tympanic bullae [GU98]; pterygoid sinuses [FB94] [GU98] partially isolated periotics [GU98]; large mandibular canals with lateral acoustic fenestrae [GU98]; cheek teeth retain complex morphology and functional occlusion [FB94] [GU98]; heavy wear facets (food was chewed) [GU98]; molars and premolars with 2 roots [C88]; loss of M3; elongate, with light, cancelous vertebrae probably filled with oil [G98] [Z+00]; short cervical vertebrae [T+01]; vertebral centra generally long and barrel-shaped [Z98]; elongated post-cervical centra [G98] [U98]; thoracic and lumbar vertebrae approximately same length [T+01]; ~15 thoracics, with last 7 lumbarized [B98]; thoracics moderately wide (width/height 1.3-1.5) [B98]; thoracic ribs strongly pachyostosed [M98]; long torso all involved in undulation [B98]; all lumbarized vertebrae elongated, with length ~twice height and width only slightly greater than height [B98]; 15 lumbars (includes "sacrals") [B98]; unfused sacral vertebrae [T+01]; sacro-illiac "joint" absent [T+01]; short tail with ball & socket vertebral articulations [T+01]; anterior 6 caudals also lumbarized [B98]; probable peduncle & tail fluke based on midcaudal reduction in centrum length & centra wider than tall [B98]; broad, fan-shaped scapulae [T+01]; scapula with anterior acromion process and small supraspinous fossa [T+01]; humerus much longer than radius [G98]; elbow joint primitively mobile [G98]; ulna with large and transversely flat olecranon [T+01]; forelimbs paddle-shaped [T+01]; forelimbs relatively narrow [Z98]; simplified pelvis joined only ventrally [Z98]; pelvis shorter than any thoracic vertebra [U98]; pelvis not attached to spine [G98]; pubis greatly elongated [U98]; pubis located anterior to obturator foramen (implying that pelvis has been rotated counterclockwise, assuming a right lateral view) [U98]; hind limbs vestigial and internal [T+01]; hind limbs could assume only two positions, used as copulatory guide? [Z98]; femurs small & gracile, shorter than innominate [U98]; femur containing extremely dense bone in central shaft (diaphyseal bone) [M98]; proximal femur sometimes malformed [U98]; femur with patellar groove [U98] patella present, ovate & thick [U98]; tibia & fibula fused [U98]; tarsals variably fused [U98]; pes I absent, pes II vestigial [U98]; may have retained terrestrial habit seasonally; long, flat plateau in vertebral morphology indicates anguilliform motion probably first cosmopolitan (not just Tethys) cetacean group [Z98]; anguilliform swimmers, using lateral undulation, who lived primarily on the surface waters [G98] (contra [B98]: anguilliform swimmers with dorsoventral undulation).
Image: Basilosaurus cetoides from Mysticetes.
Links: BBC - Beasts - Evidence - Programme 2 - Basilosaurus nice Q&A); NMNH Virtual Tour - Life in the Ancient Seas image); Primitive Eocene Whales summary of research emphasizing historical aspects); Walking With Prehistoric Beasts -- Discovery Channel -- Beast ...; Basilosaurus sp. image of fossil skull); Digital "I" Designs - Cenozoic Images - Basilosaurus Dance Basilosaurus image -- several others on this site); Basilosaurus basic information); Alabama State Fossil; WHALE FOSSILS AND EVOLUTION - Enchanted Learning Software; Archaeoceti; The Emergence of Whales, Chp. 2; Paleontology. ATW030118
References: Buchholtz 1998) [B98]; Carroll 1988) [C88]; Fordyce & Barnes (1994) [FB94]; Gingerich 1998) [G98]; Gingerich & Uhen (1998) [GU98]; Madar 1998) [M98]; Thewissen et al. (2001) [T+01]; Uhen 1998) [U98]; Zalmout et al. (2000) [Z+00]; Zimmer 1998) [Z98]. ATW030126.
 Dorudon: Gibbes 1845. D. serratus Gibbes 1845; D. atrox Andrews 1906. All other species have been referred elsewhere or synonymized.
Dorudon: Gibbes 1845. D. serratus Gibbes 1845; D. atrox Andrews 1906. All other species have been referred elsewhere or synonymized.
Range: Middle Eocene Bartonian) to Late Eocene (Priabonian) of Africa and North America (Western Atlantic & Mid-Tethys) [G98] [U98]
Phylogeny: Basilosauridae :: Autoceta + *.
Characters: about 5m long; head long (~60 cm) [V+00]; brain small; skull bones not strongly telescoped [V+00]; melon absent; complex teeth [V+00]; relatively long & flexible neck [Z98]; long & heavily muscled lumbar region; thoracic & lumbar centra not elongated [KF97]; tail pedicle not constricted [Z98]; "baseball" vertebra in tail suggest tail fluke present [Z98]; humerus much longer than radius [G98]; elbow joint mobile [G98]; forelimbs with thin cortical bone and lightweight diffuse trabecular bone [M98]; forelimbs flippered; hind limbs not in contact with pelvis (if present); complete, but small hind limbs; hind limbs & pelvis similar to other basilosaurids [Z98]; probably not efficient swimmer compared to modern whales.
Note: Uhen [U99] finds a monophyletic group containing Basilosaurus, Dorudon and Pontogeneus to the exclusion of autoctes. If correct, this may run contrary to some of the points we discuss concerning locomotion.
Links: BBC - Beasts - Evidence - Programme 2 - Dorudon; DORUDON OSIRIS Russian); Philip D. Gingerich Publications (nice cites, but also some pdf files); Research on the Origin and Early Evolution of Whales (Cetacea); Whales in the Making a b c 1. Zygorhiza , Prozeuglodon , Dorudon ....
References: Gingerich 1998) [G98]; Köhler & Fordyce (1997) [KF87]; Madar (1998) [M98]; Uhen (1999) [U99]; Vaughan et al. (2000) [V+00]; Zimmer 1998) [Z98]. ATW030126
checked ATW051030