
Reconstruction of Provelosaurus brasilienses, from Bagé-Aceguá highway, Western Rio Grande do Sul State. Length: 2.5 meters. From Some Fossil Vertebrates From Southern Brazil - Laboratory of Paleovertebrates
| Anapsida | ||
| The Vertebrates | Pareiasauridae (4) |
| Vertebrates Home | Vertebrate | Vertebrate |
|
Abreviated Dendrogram
Reptiliomorpha
├─Synapsida
└─┬─Eureptilia
│
└─Anapsida
├─Millerettidae
├─Mesosauridae
└─┬─Lanthanosuchidae
└─┬─Bolosauridae
└─Procolophonia
├─┬─Nycteroleteridae
│ └─Pareiasauridae
│ ╞═Bradysaurus
│ └─Velosauria
│ ╞═Embrithosaurus
│ ├?─Provelosaurus (biostratigraphy and large scale evolution)
│ └─┬─Deltavjatia
│ └─┬─Pareiasaurus
│ └─┬─Scutosaurus
│ └─┬─Elginia
│ └─Pumiliopareiasauria
│ ├?─Provelosaurus (morphology only)
│ └─┬─Nanopareia
│ └─┬─Anthodon
│ └─Pumilopareia
└─Procolophonoidea
├─Owenettidae
│ └?─Testudines
└─Procolophonidae
|
Contents
Overview |
Reconstruction of Provelosaurus brasilienses, from Bagé-Aceguá highway, Western Rio Grande do Sul State. Length: 2.5 meters. From Some Fossil Vertebrates From Southern Brazil - Laboratory of Paleovertebrates |
The Brazilian pareiasaur Provelosaurus americanus (Araújo, 1985), known from the nearly complete skull and postcranial elements of at least four individuals, from the Rio do Rasto Formation, Paraná Basin, Rio Grande do Sula, Brazil. Originally described as a South American representative of the genus Pareiasaurus, it was assigned to a new genus Provelosaurus by Lee (1997), who noted it shows more affinities with the small, highly derived, South African dwarf pareiasaurs (called Pumiliopareiasauria by Jalil & Janvier 2005 ) than with the more typical Pareiasaurus. According to Lee, Provelosaurus bridges the morphological gap between the advanced South African and the more generalized pareiasaurs. So far so good, African and Brazilian fauna often are very similar, look at Mesosaurus; that is just more evidence of them being adjacent pieces of the ancient supercontinent of Gondwana.
What is strange though is the date of this fauna. The Rio do Rasto Formation, which occurs at a number of different outcrops, contains a rich assemblage of classic Middle Permian tetrapods, including Platyoposaurine and Melosaurine archegosaurs and Rhinesuchoid temnospondyls among the amphibians, and such anteosaurid and titanosuchid dinocephalians and a new basal anomodont among the therapsids. Exccept fro the rhinesuchid, the otehr taxa put the minimum age limit of this fauna as mid Capitanian, and some taxa would seem to push it back to Wordian (Cisneros et al 2005, Dias-da-Silva 2012)
Yet while primitive pareiasaurs are known from the Capitanian, Provelosaurus seems anything but. It has a reduced body size, and specializations in the humerus otherwise found only in pumiliopareiasaurs and turtles, although it also retains more generalised features such as a plate-like scapula blade, unfused osteoderms, and a prominent cnemial crest. (Lee 1997 p.267). Autapomorphies in the shape of the squamosal and the morphology and placement of the quadratojugal bosses are quite unlike contemporary Tapinocephalus Assemblage Zone fauna such as Bradysaurus, Embrithosaurus, and instead show similarity with advanced forms such as Scutosaurus and Pumiliopareia (Cisneros et al 2005). A Wuchiapingian stem- pumiliopareiasaur would nicely bridge the ghost lineage and verify Lee (1997)'s and Jalil & Janvier 2005's Velosauria : Therischia + Pumiliopareiasauria hypothesis. But an early Capitanian, still less a Wordian age, Pumiliopareiasaurian makes about as much sense as a Connecitect Yankee in King Arthur's Court. Even if we assume a very early (Wordian-Capitanian) origin for a monophyletic dwarf-pareiasaur lineage, with Pumiliopareiasaurs originating in West Gondwana before migrating south-east to Africa, this does not explain why no further Pumiliopareiasaurs appear for another ten million years, in the latest Permian Latest Wuchiapingian - Changhsingian, or the otherwise sequential evolution of the pareiasaur group, nor why everything else found in the Rio do Rasto formation is very mundanely and cosmopolitanly Middle Permian, with absolutely no other evidence of endemism or precocious taxa. As with the early advanced Australian lycopod, Barragwanatha, which apparently (on the basis of associated graptolite index fossils) predates even the most primitive land plants such as Cooksonia, Provelosaurus appears to be a time travelling taxon that is much too early for its own advanced evolutionary status.
It would perhaps be too much to expect the creationists - who are so concerned to refute the fossil record that they depend on Paluxy man tracks and dinosaurs on Noah's Ark - to show any interest in rare genuine enigmas that challenge common sense by placing apparently more advanced forms earlier than more primitive ones, although non-cladistic evolutionists might, as shown by argument by critics of birds from dinosaurs used regarding the temporal paradox of Archaeopteryx occuring earlier than its "ancestor" Deinonychus; this has bnow been resolved with the discovery of maniraptoran dinosaurs older than Archeopteryx. Of course, we may actually be dealing with an authentic time travelling Anthodon, perhaps a Time Lord pareiasaur and his Tardis. Apart from this, we might consider (and/or discard) the three possible rational explanations.
(1) The least controversial possibility that Provelosaurus occurs at a latest Permian stratigraphic date (Barberena 1985) or level (Langer 2000) , as originally suggested, has now pretty much been refuted by further discoveries, which have firmly cemented the age of the tetrapod fauna as middle Permian (Malabarba et al 2003, Cisneros et al 2005, Dias-da-Silva 2012).
(2) The usual approach in current vertebrate paleontological literature is to take morphological analysis as absolute. There were pumiliopareiasaurs in the Capitanian, whether we like it or not. Almost all pareiasaur evolution is hidden from us, and can only be traced with ghost lineages, which means that ecological crises such as the late Capitanian extinction event, and any follow up adaptive radiation, were pretty insignificant, and that the fossil record is not a reliable guide to the evolutionary history of life through deep time. This phylopessimistic morphology-only approach of much of paleoherpetology-based cladistics asserts that the fossil record is so incomplete that no useful phylogenetic signal can be extracted from stratigraphy (or for that matter, geography). Cladistics still adopts a sort of atomistic idealism approach, assuming that morphological traits occur in some ideal mathematical platonic world unfettered by such mundane and messy details as stratigraphic sequence, paleogeography, and paleoecology. Moreover, the presence or absense of individual traits are considered in atomistic isolation from that of other traits, rather than considering the whole organism, its development, and its possible environment holistically. But conflicting phylogenies from molecular phylogeny alone should be enough to show that a morphology-only approach can not provide the complete story. Clearly, homoplasy and reversals play a much larger than role than is usually considered in parsimony or even maximum likelihood analyses. This is not to deny the usefulness and validity of the phylopessimistic morphology-based approach, only to suggest that it should be used as complementary to other approaches, not as the only way.
(3) It may well be that, as with the evolution of life in general, pareiasaur evolution is strongly homoplastic, and Provelosaurus developed advanced characteristics by convergence with later groups. Indeed, these convergences may simply be due to small size (suggested in passing on Palaeocritti), in the same way that lepospondyls and early amniotes appear to belong to the same clade for the same reason (Carroll, 2009). Although this is not a perfect explanation, as it does not explain the more primitive morphology of the similarily diminutive Deltavjatia, it at least has the advantage of a hypothesis in keeping with biogeographic and stratigraphic factors. Pending further developments therefore, we have decided to follow this more "holistic" approach. MAK120331

Elginia mirabilis Newton 1892
Stratigraphic range: Elgin Formation of Scotland (Dicynodon biochron ( (Lucas 2001 pp.85-7) Gordonia being a junior synonym), so Changhsingian in age)
Phylogeny: Velosauria : Embrithosaurus + (Deltavjatia + (Pareiasaurus + (Scutosaurus + (Pumiliopareiasauria + * ))))
Estimated overall size: length 60 cm long, weight about 8 or 10 kg
Characters: long conical horns over the skull and along cheek margins; notched skull table with supernumerary bones enlarged and contacting medially, forming a frill at the rear; median depression just behind the nasal bosses; posteroventral embayment of the orbital margin - Lee 1997
Comments: For a long time this was the only species of the monotypal family or subfamily Elgininiinae Cope 1895. This is a small specialised late lineage, characterised by large spikes or backwardly curved "horns". Despite the number of times it has been illustrated and referred to, Elginia is known only from a single skull, which is about 15 cm long, triangular, coarsely sculptured, and armed with a number of paired bosses or spines. The upper jaw bears 12 pairs of teeth, each with 9 or 10 cusps. The teeth are slightly constricted at the base and serrated at the crown. Zittel & Eastman 1932 p.243;
Recently, elgininiine pareiasaurs have been reported from the late Dzulfian age of Northern Pangea (Scotland and Russia). In the Moscow Basin (middle part of the Vyatskian horizon), associated with therapsids like Nanocynodon and Dicynodon, proterosuchid archosaurs, and bystrowianid amphibians, and representing a later time period than that of the giant Scutosaurs (Ivachnenko 1992). These include two partial skeletons of adults, and a young individual 25 cm long from the Vyazniki assemblage of the Vladimir region (Russian Wikipedia) A new species, Obirkovia gladiator Bulanov & Jashina 2005, was described based on a quadratojugal from the Obirkovo locality (Upper Tatarian Substage, Vologda region). Elginiids first appear in Eastern Europe in the middle of the Vyatkian Regional Stage (Scutosaurus karpinskii Zone) (Bulanov & Jashina 2005) which would be latest Wuchiapingian or Earliets Changhsingian age (Newell et al 2010, fig2 (stratigraphic correlation table)). In comparison with the earlier and abundant scutosaurs, they represent a rare element of Russian Vyatskian faunas (Golubev 2000). It may be that, as with the contemporary gondwanan pumiliopareiasaurs, these laurasian dwarf parieasaurs represent the specialised end of the line for these great herbivores.
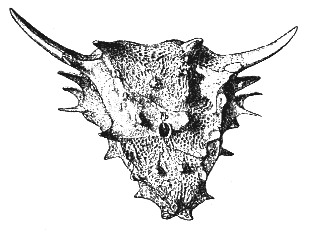 |
 Elginia mirabilis, left: illustration from Zittel & Eastman 1932 p.242, right, side view, sketch from Kuhn, 1969, p.74 |
As with many pareiasaurs, precise phylogenetic placement is uncertain. Elginia shares with Scutosaurus elaborate cranial ornament, which may imply they are related (Boonstra, 1932), although this is more likely to be the result of simple convergence. It was also thought related to the very primitive taxon 'Dolichopareia' (=Nochelesaurus) on the basis of a deeply notched skull table shared with the latter (Walker 1973) , but this turned out to be the result of taphonomic distortion (Lee 1997). Cladistic analyses tend to place it as more derived than the earlier giant Pareiasaurus and Scutosaurus, but evidence of actual phylogenetic relationships is obscure, and phylogenies differ according to whether Elginia is the most highly derived Therischian ( Lee 1997, Jalil & Janvier 2005) or intermediate between Scutosaurus and Pumiliopareiasaurs (Tsuji & Müller 2009).
Links more on Elginia
Image Elginia life reconstruction, artwork by Dmitry Bogdanov, Wikipedia, public domain.
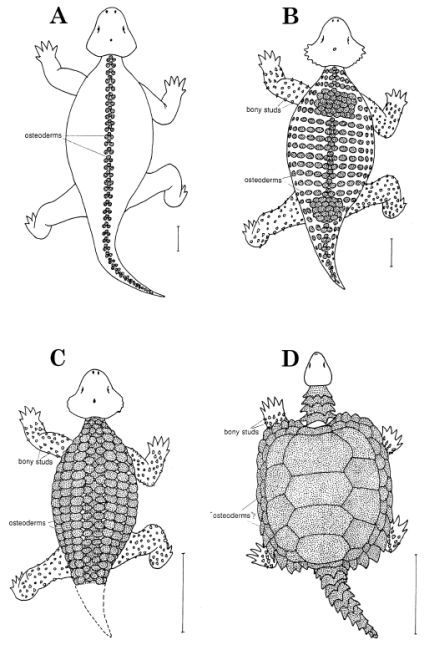 Pareiasaurs and turtles - from Lee 1997 fig.19. showing dorsal dermal armour and bony studs over limbs, and overall similarity in body proportions. Dorsal reconstructions of (A) the large primitive pareiasaur Bradysaurus (Middle Permian of South Africa), (B) the large advanced paraeiasaur Scutosaurus (Late Permian of Russia), (C) the small advanced pareiasaur Anthodon (Latest Permian of South Africa), and (D) the stem chelonian Proganochelys (Late Triassic of Germany). Scale bar 30 cm |
Pumiliopareiasauria Jalil and Janvier 2005 : Anthodon, Nanoparia, and Pumiliopareia
Range: Latest Permian of Sth Afr
Phylogeny: Velosauria : Embrithosaurus + (Deltavjatia + (Pareiasaurus + (Scutosaurus + (Elginia + * : Nanopareia + (Anthodon + Pumilopareia)))))
Comments: [1] These are the dwarf pareiasaurs, the most highly derived, the heavily armoured, and turtle-like of the group, that only appear at the end of the Permian. Anthodon and Pumiliopareia share numerous and striking postcranial traits with turtles, these would be synapomorphies if they are turtle ancestors (Lee 1997, pp. 267-8) and convergences if they are not
[2] The idea that turtles are actually highly derived pareiasaurs was strongly argued on cladistic grounds by Michael Lee in a series of innovative papers (Lee, 1993, 1995, 1997, 2001 ), and supported by Jalil & Janvier 2005. Thus, pareiasaurs had turtle-like cranial features, and it is not hard to imagine dermal ossicles and scutes, embedde din the skin or attached to the skull, vertebrae, progressively evolving into bony plates,and finally fusing into a turtle shell. There is also a progressive morphological sequence, as illustrated in the diagram at the right, showing pareiasaurs becoming progressively more heavily armoured and then, after a series of giant forms, progressively smaller. Pareiasaurs evolving into turtles also has a coolness factor rather like dinosaurs evolved into birds. Significantly, both came about through the new science of cladistics. It would also mean, cladistically speaking, that turtles are pareiasaurs, although surely this is no worse than alternatives such as turtles are plesiosaurs! (well, sauropterygians, but let's not quibble!) But whereas dinosaurs to birds is as solid a phylogenetic hypothesis as any that could be formulated, the question of a pareiasaur origin for turtles and tortoises is a lot more controversial, and, unlike dinosaurs to birds, there are some strong rivals. deBraga & Rieppel, 1997 argue that pareiasaur scutes are not homologous with the turtle shell and show that many features of turtles, especially the limbs, suggest sauropterygia and lepidosauromorph relationships. Meanwhile molecular phylogeny consistently places turtles among the archosaurs. Recently the eunotosaur ancestry hypothesis has been revived. None of these hypotheses is without controversy, but if pareiasaurs are not rturtle ancestors, than irather than evolving into turtles, pareiasaurs were evolving parallel to them. As such, pareiasaurs would represent "prophetic forms" rather than true ancestors. MAK120331
[3] The middle Permian Provelosaurus americanus is commonly placed as the most basal pumiliopareiasaur, but stratigraphically this makes no sense, see our comments above
[4] on a much more trivial level, here at Palaeos we need to reconsider our colour attribution; having orange for extinct clade sor taxa and yellow for extant ones is not very useful if it is not even clear whether a clade is actuallyextinct or extant or extant. If MSY Lee is correct then all the dendrogram links and headings dfeaturing pareiasaur clades need to be change dfrom orange to yellow (and otehrs such as procolophonidea etc from yellow to orange) . One controversy-proof option (albeit a bit colourful) would be to introduce a colour coding based on earliest appearance of the taxon in question a la the geological timescale; but to change everything would be so much work, given thousand and thousand of entries, it is not presently considered practical MAK120402
Nanoparia luckhoffi Broom 1936
Stratigraphic range: Dicynodon (previously called Daptocephalus) Zone, Lower Beaufort Beds, Karoo basin, South Africa (Changhsingian age)
Phylogeny: Pumiliopareiasauria : Nanopareia + (Anthodon + Pumilopareia)
Size: length 60 cm, weight about 8 or 10 kg
Characters: snout very pointed in dorsal view, superficial element present between the squamosal and quadratojugal. Lee 1997, p. 210
Comments: This is an unusual small, spiny specialised form. The skull is very similar to that of Pareiasaurus and Romer considered it a synonym of the latter. Orlov however (in Osnovy Paleontology, the monumental multi-volume Russian textbook of Paleontology) placed it in the Elginiinae. Kuhn 1969 however argues that while resembling Elginia in the ossifications at the rear of the skull, it differs completely in proportions and would not seem to be related. Lee 1997 considers it a basal member of the dwarf pareiasaurs.

Anthodon serrarius Owen 1876
Synonyms: Propappus parvus Haughton, 1913, Pareiasaurus parvus (Haughton, 1913)
Stratigraphic range and localities: late Early to early Late Dicynodon (=Daptocephalus) zone (Ward et al 2005, supplem. info, range data; ), Lower Beaufort Beds, Karoo basin, (latest Wuchiapingian to Middle Changhsingian age)
Phylogeny: Pumiliopareiasauria : Nanopareia + (Anthodon + Pumilopareia)
Size: Skull length 30 cm, overall length 1.2-1.5 m long, estimated weight about 80 to 100 kg
Characters: marginal teeth do not form a continuous cutting edge, but separated by small gaps; the cranial sculpture consists of fine, widely-spaced pits and weakly developed bosses. - Lee 1997, p. 210
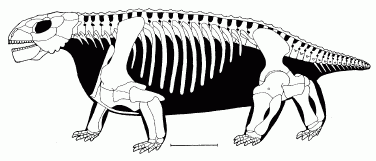
Comments: this small form combines the primitive feature of interpterygoid fenestra with an advanced feature of turtle-like armour. The skull is small and quite lightly built. The cheekbones form very large quadratojugal "horns" that extend downwards to a great degree, but with a smooth unornamented surface. The mandible has ventral protrusions (further "horns"). The postparietals are fused and, along with the tabulars, located on the skull roof, as in more primitive diadectomorphs. Small dermal ossicles covered the body, while the pattern of armour plates on the back reminiscent of a turtle shell. There are 11 to 14 pairs of overlapping teeth, of small and uniform size, each with 8 to 15 cusps, giving them, as with all pareiasaurs, a leaf-like or flower like appearance, hence the genric name "flower tooth". The tail was further shortened relative to less derived forms.
The Holotype (BP/1/548) consists of an incomplete skull and several anterior vertebrae. The teeth are regularly spaced and have at least 9 cusps each.
Anthodon minusculus Haughton 1932 from the Daptocephalus zone, Ruhuhuh valley, Tanzania is known from a single humerus and would be a nomen dubium
Graphics Sketch of skull from from Kuhn, 1969 Skeletal reconstruction from Lee 1997
Links Palaeocritti - Anthodon serrarius, David Peters - Anthodon,
Pumiliopareia pricei (Broom and Robinson 1948)
Synonyms: Nanoparia pricei Broom and Robinson 1948, Pareiasaurus pricei (Broom & Robinson, 1948),
Range: Dicynodon zone of Sth Afr (Changhsingian)
Phylogeny: Pumiliopareiasauria : Nanopareia + (Anthodon + Pumilopareia)
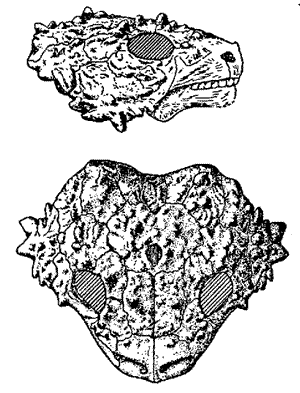
Size: Length of skull 12 cm; overall length about 50 cm.Characters: skull table and cheeks completely covered by densely-spaced, conical bosses; orbits perfectly round; only approximately 9 teeth on each upper jaw and 7 on each lower jaw; ribs very wide, abutting, the intercostal spaces thus being almost non-existent. Lee 1997, p. 210
Comments: Known from a complete skeleton with osteoderms, Pumiliopareia is the smallest known member of the pareiasaurs, measuring only a fifth the linear dimensions of its larger relatives. Like Anthodon, its body was entirely covered with osteoderms. In analyses that support a pareiasaur origin of turtles, the sister taxon of the testudines. However it specifically shares with shares with turtles a single triat only: Ribs greatly expanded anteroposteriorly (i.e. wide) Originally included under the genus Nanoparia, it was given its own name by Lee 1997 who found it did not form a clade with Nanoparia luckhoffi, the type species of that genus, and preferred to have monophyletic genera. Nanoparia may still be a paraphyletic genus, which is allowed in Linanean binomial taxonomy, or it may be that all three Pumiliopareiasaurs are similar enough to belong to single genus. For now we have retained Lee's monotypal genus; the name, taken from the Latin pumilio - dwarf , appropriately refers to the diminutive size of these animals. Lee 1997 p.268 MAK120401
Links Palaeocritti - Pumiliopareia pricei
Graphics: Illustration of skull from Kuhn, 1969, p.79
page MAK120322