
| Anapsida | ||
| The Vertebrates | Pareiasauridae (2) |
| Vertebrates Home | Vertebrate | Vertebrate |
Velosauria : Anthodon, Deltavjatia, Pareiasuchus, Scutosaurus. Typical pareiasaurs.
Middle to Late Permian
Pareiasauridae : Bradysaurus + * : Embrithosaurus + (Deltavjatia + ((Pareiasaurus + (Scutosaurus + (Elginia + Pumiliopareiasauria))))
$ 12 or more cusps on teeth of lower jaw; $ parallel, closely spaced rows of palatal teeth; $ parasphenoid greatly shortened; large, U-shaped interpterygoid vacuity; $ <20 caudal vertebrae; cleithrum absent; $ greater trochanter present and deflected (?) from femoral head; $ bony dermal armor, with large osteoderms; $ armor covers entire dorsal surface, united over shoulder and pelvis; $ conical studs over appendages.
References: deBraga & Rieppel (1997); Lee (1997). ATW020924
Links: Introduction to Pareiasauria; Parareptilia; Scutosaurus karpinski.
Images: Scutosaurus karpinskii © mathematical.com, used with permission. ATW020924. revised MAK120331
Comments: (1) Includes the Subfamily Pareiasaurinae Nopcsa 1923. Oskar Kuhn (Cotylosauria, p.75) includes the following genera here: Pareiasaurus, Pareiasuchus (Dr Carroll includes this genus under Pareiasaurus), Embrithosaurus, Anthodon, Scutosaurus, Shihtienfenia, and Nanoparia. Von Huene however places Scutosaurus, Shihtienfenia, and Nanoparia each in their own subfamilies. Cladistics substituted taxonomy with phylogeny, but this didn't help because hypotheses keep changing. Lee (1997) argues that this group (his clade 'D') is paraphyletic and should include Testudines (or Chelonia). The current consensus is that the pareiasaur origin is no longer valid, the two most popular turtle origin hypotheses now being thequite morphologically implausible crown archosauria of molecular phylogeny, and sauropterygian lepidosauromorpha, although we have gone for a more conservative prtocolophonoid origin
(2) includes the "typical" Pareiasaurs. The heads decorated by knobs and ridges. The teeth each have nine cusps. The upper margin of the ilium is flat, never vaulted. The surface of the back is armoured to varying degrees with bony scutes. Some forms like the ancestral genus Embrithosaurus had only weak armour; while later forms like Pareiasaurus, Scutosaurus, and Anthodon were well protected. The limb posture is advanced and upright to support the weight of the body (graviportal).
(3)In view of the paraphyletic status of Bradysaurus (Tsuji & Müller 2009) there doesn't seem to be a clear point where node Velosauria should be placed. Mikko Haraama offers two alternatives, Velosauria Lee, 1994 sensu Lee, 1997, which is an apomorphy-based definition, everything more derived than Bradysaurus, and Velosauria Lee, 1994 sensu Jalil & Janvier, 2005, which is a node-based definition (Therischia + Pumiliopareiasauria) for everything more derived than Deltavjatia. Given that there is no unanimity regarding the Therischia - Tsuji & Müller 2009's unpublished analysis (see cladogram in that paper) did not recover it - we have gone for the former option. MAK120323
Embrithosaurus Watson 1914
Range: Mid Permian of Sth Afr
Phylogeny: Velosauria : Embrithosaurus (Nochelesaurus) alexanderi + (Embrithosaurus schwarzi + (Deltavjatia + (Pareiasaurus + (Scutosaurus + (Elginia + Pumiliopareiasauria)))))
synonyms: Dolichopareia, Nochelosaurus
Stratigraphic range: Lower to Upper Tapinocephalus zone, Lower Beaufort Beds, of the Karoo basin, South Africa; most common in the Lower Tapinocephalus zone (Lower to Middle Tapinocephalus zone in Boonstra, 1969 pp26, 32,; Lower Tapinocephalus to Pristerognathus (= old Upper Tapinocephalus) zone in Ward et al 2005, supplem. info, range data; both = Capitanian age)
length: about 2.5 meters long
weight: about 600 kg
Comments: [1] Embrithosaurus is another huge early Pareiasaur, a contemporary of Bradysaurus. The skull is relatively deep and narrow. The body is lightly armoured with thin, smooth dermal scutes. This genus would seem to be ancestral to Pareiasaurus. MAK000124.
[2] Lee 1995 and Jalil & Janvier 2005 have three species, and Tsuji & Müller 2009 four as a scalloped paraphyletic series. Four pareiasaur taxa (Bradysaurus baini, Bradysaurus seeleyi, Embrithosaurus (Nochelesaurus) alexanderi and Embrithosaurus schwarzi co-existing in the same environment seem excessive, so either this was intraspecies variation, gender difference, a stratigraphic and anagenetic (chronospecies) series, or niche partitioning MAK120326
[3] Cladistic analysis makes Embrithosaurus as originally described paraphyletic, or in other words, an evolutionarily intermediate form between Bradysaurus and higher pareiasaurs, and to avoid paraphyletic taxa the synonym Nochelosaurus is restored. The present writer (MAK) is opposed to the prohibition against representing transitional and ancestral species in accounts of Earth History, as he feels that this is a too literal application of cladistic formalism, which is concerned with hypothesis testing, not with describing the actual evolutionary history of species in deep time. Also, although phylogenetic nomenclature tends towards monospecific genera (hence preference here of Nochelesaurus over Embrithosaurus, this creates an incongruency with taxonomy of plants, invertebrates, and quaternary and recent vertebrates, all of which ares based multi-species genera. For this reason, Embrithosaurus is retained for all species in these evolutionary series (ditto for Bradysaurus and Pareiasaurus); in any case there is no restriction on paraphyletic genera in linnaean (binomial) taxonomy. MAK120329
 
|
Embrithosaurus strubeni (Broom, 1924)
Synonyms: Bradysaurus strubeni in Boonstra, 1969 p.30
Stratigraphic range: Lower to Upper Tapinocephalus zone, Lower Beaufort Beds, South Africa
Comments:The skull is large and deep, pointed at the front, and elevated in the jugal region. This species was originally made the type species of Nochelosaurus by Haughton and Boonstra. Boonstra later (1969) moved it into the genus Bradysaurus, on the basis of the primitive tooth structure. Kuhn however considers it belongs under Embrithosaurus
The specimen shown in lateral view (top) is 48 cm long, the other 35 cm long MAK000124
 
|
Embrithosaurus alexanderi (Haughton and Boonstra 1929)
Synonyms: Nochelesaurus angustus Haughton and Boonstra, 1929, Embrithosaurus angustus (Haughton and Boonstra, 1929), Dolichopareia angusta Haughton and Boonstra, 1929, Brachypareia watsoni Haughton and Boonstra, 1929, Pareiasaurus bombidens Seeley, 1888, Bradysaurus watsoni (Haughton & Boonstra, 1929)
Stratigraphic range: Lower to Upper Tapinocephalus zone, Lower Beaufort Beds, South Africa
Size: Skull Length about 55 cm, Overall length about 3 m
Comments: This species was made the type for " Dolichopareia". As the name indicates, the skull is long and narrow. This would seem to indicate a different lifestyle or diet to other parieasaurs. More recently, it has been used as the monotypal species for the genus Nochelesaurus (it is not clear what the status of Embrithosaurus strubeni is, this may be a further transitional species) The bottom skull is about 48 cm long, and was originally described as Brachypareia watsoni, In cladistic analyses, this species is phylogenetically intermediate between Bradysaurus seeleyi and Embrithosaurus schwarzi MAK000124 120329
Graphics sketches of skulls of Embrithosaurus angustus (top) and Bradysaurus watsoni (bottom). These two species are considered synonyms of Embrithosaurus alexanderi, which indicates either artistic license or a great deal of individual variation, postmortem distortion, and so on. From Kuhn 1969 MAK120401
 |
Embrithosaurus schwarzi Watson 1914
Stratigraphic range: Lower to Upper Tapinocephalus zone
Size: Overall length: 2.2 m
Characters: The bony dermal plates are thin and smooth. The ulna is without an olecranon process. (Zittel & Eastman 1932 p.243) MAK000124. "Anterior expansion of the iliac blade is flat rather than everted; the two iliac blades are not parallel but diverge anteriorly, making an angle of approximately 40° with the sagittal plane; pelvic symphysis is extremely thick, being almost half as deep as long." - Lee 1997
Comments: The type species. This is the most advanced species of this genus, as indicated by the teeth, which have nine cusps (in three groups of three). In cladistic analyses it is used as the monotypal species for the genus.
Graphics illustration of skull from Kuhn 1969

Deltavjatia vjatkensis (Hartmann-Weinberg 1937)
Synonyms: Anthodon chlynoviensis Efremov, 1933, Anthodon rossicus Hartmann-Weinberg, 1933, Pareiasuchus vjatkensis Hartmann-Weinberg, 1933, Scutosaurus rossicus (Hartmann-Weinberg, 1933)
Range: Severondvinskian Gorizont (= Lower Zone IV, Tatarian stage), Late Capitanian, (Latest Middle Permian ) Kotel'nich locality in central Russia
Phylogeny: Velosauria : Embrithosaurus (Nochelesaurus) alexanderi + (Embrithosaurus schwarzi + ((Pareiasaurus + (Scutosaurus + (Elginia + Pumiliopareiasauria))) + *)
Size: 1 to 2 meters long
Characters: "the ventrolateral cheek flange terminates in a long, flattened, rectangular boss; the occipital condyle projects far behind the posterior border of the postparietal; in the largest specimen, the boss on the postfrontal is enlarged into a long conical horn" Lee 1997
Comments: Originally placed in Pareiasuchus (Hartmann-Weinberg 1933) this smallish pareiasaur is distinct enough to warrant its own genus. The only pareiasaur known from the lower Tatarian of Russia, Deltavjatia is an important taxon that bridges the large morphological and stratigraphic gap separating primitive South African Tapinocephalus zone pareiasaurs from the more derived forms characteristic of the overlying Cistecephalus zone and later deposits. It retains the long tail and feeble dermal armour of more primitive pareiasaurs, but possesses many skeletal traits characteristic of more advanced species Lee 1997. From above, the skull resembles the letter V, hence the name Deltavjatia (from the Greek letter Delta).
In spite of the large eye sockets, the eyeballs were small, as is shown by the size of the sclerotic ring that reinforced the cornea. Skulls of pareiasaurs with the sclerotic ring preserved are very rare. As with all pareiasaurs, the surface of skull is uneven. being covered with horn-like bumps and ossicles (bony plates) of different sizes. The largest of these are usually on the cheek, parietal and nosal bones. Sometimes there are large bosses on the frontal bones, and almost always on the lower jaw where it articulates with the skull. Attached to the largest dermal ossicles are nodules or bosses of various sizes. The spaces between the dermal ossicles probably contained skin glands, which exuded specific muscus for keeping the skin moist, or even, as with some toads, poisonous warts to deter predators. (Vyatka Palaeontological Museum)
As with all pareiasaurs, the skull does not contain a separation of mouth and nose cavities. The animals had to bolt their food without chewing, because when the food was in mouth cavity, there was impossible to breathe. (Vyatka Palaeontological Museum)
These animals were probably highly gregarious, occuring in large numbers. As with many of the similarily abundant South African Bradysaurs, most of the Deltavjatia found at the Kotelnich fossil locality were recovered as complete skeletons found fossilised in standing posture. The usual explanation is that they became bogged in mud died as a result of heavy rain fall, and were buried by silt and sediments carried by the flood water. Another explanation is that these water-loving animals were crowded in small ponds when there was not enough water to swim through but the ground became series of mud traps. The deltavjatias preserved this way show not only the smallest bones of the feet and tail, but also the dermal ossicles. (Vyatka Palaeontological Museum)
No doubt because of its primitive nature relative to the later and more advanced Scutosaurus, Golubev 2000 places this genus in a stratigraphic table of Russian Permian tetrapod families under the "Bradysauridae". Deltavjatia is also more lightly built than Scutosaurus, and may have been a more active animal. Despite its primitive status, it is still characterised by numerous turtle-like skull features (e.g., a very high palate), limbs, and girdles, and lateral projections flaring out some of the vertebrate in a very shell-like way. (Transitional Vertebrate Fossils FAQ by Kathleen Hunt); This relatively small species lived immediately after the mid Capitanian extinction event and represented the start of a new paraeiasaur adaptive radiation MAK120331
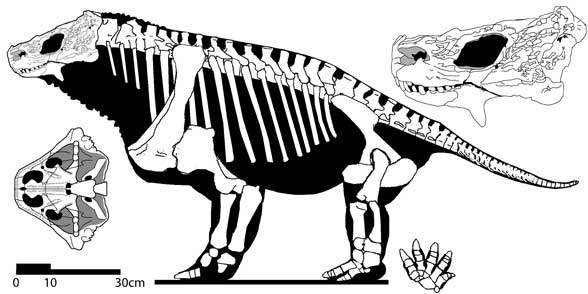
Graphics Photograph of Deltavjatia vjatkensis © Albert Khlyupin and Vyatka Palaeontological Museum - original page (Parareptilia), used with permission. Skeletal reconstruction © David Peters
page MAK120322