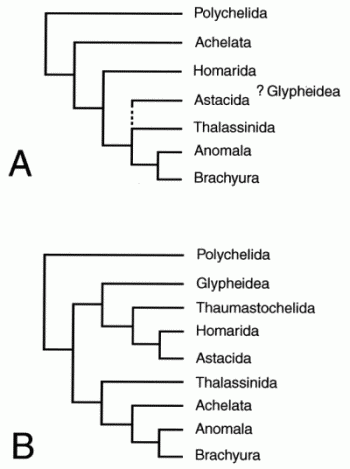
Cladistic relationships of major reptant clades based on cladistic morphology-only analyses: A, Scholtz & Richter 1995, and B, Dixon et al 2003. There is agreement at the base and top of the tree but not the middle. From Ahyong & O'Meally 2004 fig 1
| Decapoda | ||
| Arthropoda | Reptantia |
| Arthropoda | Decapoda
└─► |
|||
| Malacostraca | None |
Hexapoda |
|
Abbreviated Dendrogram
Malacostraca
│
└─Decapoda
├─Dendrobranchiata
└─┬─Caridea
└─┬─Stenopodidea
└─Reptantia
├─Palaeopalaemon
├─Eryonoidea
╘═╤═Glypheoidea
├─Astacidea
├─Thalassinidea
├─Palinuroidea
└─┬─Anomura
└─Brachyura
|
Contents
Overview |
Taxa on This Page
The Reptantia or "walkers" (reptants for short) are the clade of crustaceans containing all decapods apart from the Natantia or "swimmers" , the shrimps and prawns, (presumably these would be called natants for short, but no-one uses the term Natantia anymore). Reptantia includes crabs and crab-like forms, lobsters and lobster-like animals, and freshwater crayfish (which are related to the lobsters proper) . Reptants are larger and more heavily built, with stronger and more robust legs, than natants, as a result of the shift in ecomorph from swimming and nektonic types to bottom dwellers. They are also much more morphologically diverse, featuring seemingly every possible morphotype within the basic bauplan - large claws, small claws, no claws, one set of claws, multiple pairs of claws, both main claws of the same size, one claw much bigger than the other, antennae shorter, antennae very long, body and thorax long, body ooval shaped, body short and broad, eyes stalked, eyes not stalked, and so on. The most successful of all the reptants are the short bodied Brachyura or true crabs, which invaded the littoral zone, and account for almost half of decapod biodiversity MAK120530
|
Cladistic relationships of major reptant clades based on cladistic morphology-only analyses: A, Scholtz & Richter 1995, and B, Dixon et al 2003. There is agreement at the base and top of the tree but not the middle. From Ahyong & O'Meally 2004 fig 1 |
The uncertainity regarding reptant phylogeny is nothing unexpected; it is simply representative of the uncertainty regarding Crustacean phylogeny as a whole. There have to date been only two independent morphology-based cladistic analyses of the decapoda as a whole, those of Scholtz & Richter in 1995 and Dixon et al in 2003 (right). All agree regarding the basal (bottom of the tree) position of the Polychelida (in these pages we have used the more traditional term Eryonoidea, although Polychelidae can be applied to relict extant taxa), the polyphyly of the traditional Palinura (lobsters and lobster like forms, we use the term in a more specific context to refer to the spiny lobster Palinurus and related lineages) and a monophyletic crown clade Meiura (Anomura (crab-like forms) + Brachyura (true crabs)) as the most derived taxon (at the top of the decapod tree). Beyond that, there is no agreement. Both papers introduce a number of new clades, most of which are invalidated by the findings of the other paper; only Reptantia, Eureptantia and Meiura are recognised by both. For example the Achelata or spiny lobster group (= Palinura sensu stricto, or Palinuroidea) are either the second most basal group or the sister group of the highest clade (Meiura). There is a vague sort of consensus with the Thalassinidea, a small group previously placed in the Anomura, which is here either a possible sister group of the Meiura, or of Meiura + Achelata. A "total evidence" analysis by Ahyong & O'Meally 2004 incorporates aspects of both papers but again only the base and crown of the tree are in common with both of the other two. The problem with cladistics, despite its early promise, is that it faces the same homology vs homoplasy problem that defeated its fellow methodology phenetics. But pure molecular sequencing is no help here either. Unlike the mammals, where a quite stable consensus has emerged, with the decapods every study gives totally different results. Nor do even one of these studies match any of the rival morphology studies. For example the recent paper by Bracken et al 2009 replaces the Polychelidae with the Thalassinidea as the most he basal group. This is simply be due to anthropocentric bias - furry cuddly mammals are more like us than lobsters are - and hence there is more work done on, are more papers written about, mammals than there are about crusty crustaceans. And certainly there is more molecular consensus regarding arthropods as a whole than there is with particular groups. The problem remains though, as it does with mammals, or turtles, or any other sampled group, of the incompatability between molecules and morphology, and the morphological absurdities that a molecules-only phylogeny demands. The current tendency in phylogeny in general is certainly very much away from morphology and in favour of molecules, which seem far less suited to mapping deep phylogeny in groups that emerged through a sudden adaptive radiation over a short span of time, there now seems to be a counter movement back to very obscure neontological morpohological details and to paleontology to supplement the molecular sequencing and hopefully provoide a better picture.
The following syncretic dendrogram is abbreviated and slightly modified from the one by Christopher Taylor - Variety of LIfe and is mostly inspired by the more recent of the two cladistic studies mentioned earlier; Dixon et al 2003 ([DAS03] in the square brackets). It is provisionally included here to make it look like we know the details of the evolutionary tree, which we most patently do not. But don't use that last phrase as an excuse for creationism. Evolution is a fact, like heliocentricism, electromagnetism, and gravity. And while it can also be said with equal certainty that God, or the devil (dependng on your theology), didn't put fossils in the ground to make the Earth look old, he certainly is in the details. So, on with the show (or just skip the following if it's too boring). MAK120529
References
[B95] Bousfield, E. L. 1995. A contribution to the natural classification of Lower and Middle Cambrian arthropods: Food-gathering and feeding mechanisms. Amphipacifica 2: 3-34.
[B06] Boyko, C. B. 2006. New and historical records of polychelid lobsters (Crustacea: Decapoda: Polychelidae) from the Yale Peabody Museum collections. Bulletin of the Peabody Museum of Natural History 47 (1-2): 37-46.
[DAS03] Dixon, C. J., S. T. Ahyong & F. R. Schram. 2003. A new hypothesis of decapod phylogeny. Crustaceana 76: 935-975.
[G75] Grant, E. M. 1975. Guide to Fishes. The Co-ordinator-General's Department: Brisbane (Australia).
[H47] Hatch, M. H. 1947. The Chelifera and Isopoda of Washington and adjacent regions. University of Washington Publications in Biology 10 (5): 155-274.
[HS01] Hayward, B. W., A. B. Stephenson, M. S. Morley, W. M. Blom, H. R. Grenfell, F. J. Brook, J. L. Riley, F. Thompson & J. J. Hayward. 2001. Marine biota of Parengarenga Harbour, Northland, New Zealand. Records of the Auckland Museum 37: 45-80.
[MT08] Magalhães, C., & M. Türkay. 2008. A new species of Kingsleya from the Yanomami Indians area in the upper Rio Orinoco, Venezuela (Crustacea, Decapoda, Brachyura, Pseudothelphusidae). Senckenbergiana Biologica 88 (2): 231-237.
[N24] Nicholls, G. E. 1924. Neoniphargus branchialis, a new freshwater amphipod from south-western Australia. Journal of the Royal Society of Western Australia 10 (14): 105-111.
[PH03] Pinheiro, M. A. A., & G. Y. Hattori. 2003. Embryology of the mangrove crab Ucides cordatus (Brachyura: Ocypodidae). Journal of Crustacean Biology 23 (3): 729-737.
[S77] Schram, F. R. 1977. Paleozoogeography of Late Paleozoic and Triassic Malacostraca. Systematic Zoology 26 (4): 367-379.
[W08] Wilson, B. 2008. Background information on Faure Island, Shark Bay, Western Australia. Records of the Western Australian Museum Supplement 75: 1-9.
Fossil record only from the Mid Cretaceous; ghost lineage suggests Devonian origin
Phylogeny: Pleocyemata : Caridea + (Reptantia + * )
Comments: Stenopodidea (which contains the so-called "boxer shrimp" or "barber-pole shrimp") MAK120518
From the Devonian
Phylogeny: Pleocyemata : Caridea + (Stenopodidea + * : Palaeopalaemon + (Eryonoidea + Eureptantia)
Characters:. Synapomorphies (from Dixon et al. 2003): Pereiopods articulating in a medio-lateral manner (enabling walking); first two segments of PI fused together. Carapace is calcified and flattened, telson lacking spines. Mandible with long articulation along anterior edge. Third maxilliped shortened relative to other thoracopods; first pereiopod enlarged.. - CKT.
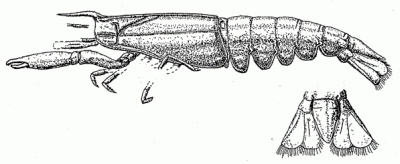
Phylogeny: Reptantia : (Eryonoidea + Eureptantia) + *
Comments: this species has been widely assigned to various repantian taxa, but about all that can be said odf it for certain is that it is the earliest known decapod. It shows that as early as the Devonnian, the decapods had already diverged into shrimp- and lobster-like lineages. Although Palaeopalaemon shares a characteristic of the Thalassinidea and the Meiura, we propose a new name, Lineata, carapace lineae, which suggests it may be related to these taxa. However this would require a number of long ghost lineages to explain the anbsense of Paleozoic fossils (Schram & Dixon, 2003). It also does not explain the poor molecular resolution of the Reptantia as a whole, which only makes sense if all the major clades in the group diverged in a short period. . We believe it is more parsimonious therefore to assume that this feature is teh result of convrgence, and appeared independently in at least three groups, and that the the Reptantia was arather insignificant and not very diverse group for the first third of its history. It was only in surviving the great P-T extinction event that Palaeopalaemon's descendants would initate perhaps the most dramatic and important event in crustacean evolution since the Cambrian explosion, the sudden and radical adaptive radiation of "crawlers" (Repantia) to replace the mass extinction of almost all other large aquatic arthropods at the end of the Paleozoic. MAK120528
Image: Palaeopalaemon newberryi, from Schram 2009
Page MAK120518