
Hermit crab
Photo by Stephen Gibson, Creative Commons Attribution.
| Decapoda | ||
| Arthropoda | Anomura |
| Arthropoda | Decapoda
└─► |
|||
| Malacostraca | None |
Hexapoda |
|
Abbreviated Dendrogram
Malacostraca
│
└─Decapoda
├─Dendrobranchiata
└─┬─Caridea
└─Reptantia
├─Eryonoidea
└─┬─Eureptantia
├─Palinuroidea
└─Meiura
├─Anomura
│ ├─Hippoidea
│ └─Aeglidae
│ ├─Galatheoidea
│ ├─Kiwa
│ ├─Lomis
│ └─┬─Paguroidea
│ └─Lithodoidea
└─Brachyura
|
Contents
Overview |
Taxa on This Page
Hermit crab Photo by Stephen Gibson, Creative Commons Attribution. |
The Anomura is a group that includes various organisms commonly called crabs, such as hermit crabs, stone crabs, mole crabs, and half crabs, that are not actually true crabs (Brachyura) . After the tryue crabs, they are the most speciose decapod group.
A morphologically diverse group, some anomurans are little different from lobsters, whereas others are very crab like in appearance. The various lineages independently evolved crab-like charcetristics. The differ from true crabs in that the abdomen is not as small. In true crabs the abdomen is tiny and folded under the thorax, to shift the center of gravity forward and improive mobility. In crab-like anomurans the abdomen may be soft or bent upon itself with reduced side-plates and tail-fan (Zittel & Eastman 1913) but it is still reasonably large, and hence anomurans are slower moving than their fast side-scuttling cousins.
The only two comprehensive cladistic analyses of the decapoda (Scholtz & Richter 1995 and Dixon et al 2003) both place anomurans as the sister group to the true crabs, the two together being known as the Meiura. Molecular analysis doesn't support this, and instead have the two groups popping up all over the Reptantian family tree without rhyme or reason (see comparison of cladograms). It seems likely that the anomura and the brachyura diverged very soon after evolving from a lobster-like ancestor; in too short a time scale to be picked up by molecular phylogeny.
As for the relations of the Anomuran subgroups (which themselves diverged somewhat later as they are not jumbled in molecular phylogeny with other decapod clades) there is no real consensus. There isn't even agreement over what the group as a whole should be called. Cladists tend to prefer Anomala, because the original definition of Anomura is parphyletic or polyphykletic as it includes the Thalassinida (which may or may not still be related). The majority of zoologists still use the Anomura, which has taxonomic priority as the older term. We have already given our opinion why excluding groups from a taxon is no reason to change the taxon's name, if the original type species is still included or the original definition still stands. For this reason the older and more widely used name of Anomura is here retained. MAK120529
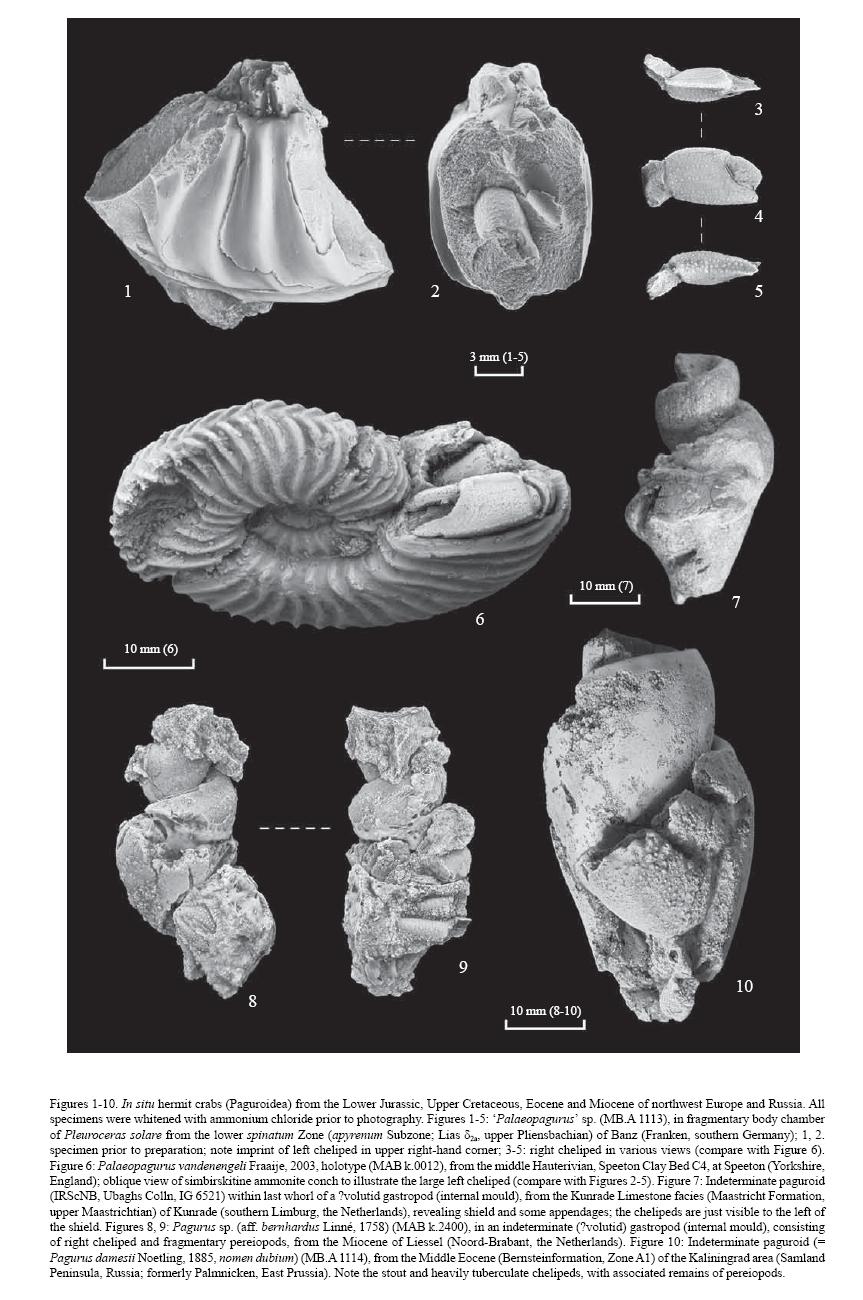
The Anomura are rare as fossils, although that is now changing. The Galatheidea (porcellan crabs) have generally been represented only by chelae (originally known only from the late Cretaceous of Denmark, and referred to the Recent Galathea). However four genera in each of the superfamilies Galatheoidea and Paguroidea were cited by Glaessner 1969 (Treatise on Invertebrate Palaeontology) as occurring as early as the Jurassic, while a review by Schweitzer & Feldmann (2000) has eight galatheoid genera with fossil records dating back to the Middle or Late Jurassic. The freshwater Aeglidae, is now known from Late Cretaceous marine rocks in New Zealand, suggesting that its freshwater adaption might be a recent development, and then discovered in the Early Cretaceous of Mexico, implying much greater distribution during the Mesozoic than at present. The Hippidea, which previously lacked a fossil record, have been reported from the Middle and Late Eocene. The Paguroidea, including the Hermit-crabs and their allies, was at first very doubtfully represented in the Eocene of Hungary by chelae referred to the Recent Pagurus. For a long time they were known in the fossil record primarily from isolated claws (chelipeds), which is not sufficent for them to be placed phylogenetically. A few descriptions of in situ gastropod shell-inhabiting hermit crabs are known from the Late Cretaceous, Oligocene, and Miocene. But a perfectly preserved Early Cretaceous hermit crab found occupying the shell of an ammonite cephalopod Fraaije (2003), apparently a diogenid from the larger left cheliped. might explain the rarity or absence of hermit crabs in gastropod shells of Jurassic and Early Cretaceous ages, if paleontologists have focused their attentions on the wrong class of molluscs. Since then, more discoveries of hermit crabs occupying ammonite and gastropod shells have been made (Jagt et al 2006, and see graphic, below). Even though hermit crabs occupied Mesozoic gastropod shells, the shift in prefernce from ammonites to gastropod shells was the logical conseque of the former's extinction at the end Mesozoic. Bakel et al. (2008) reported the discovery of a series of Jurassic age paguroid carapaces, including two that appear assignable to families of the Pylochelidae. - (Zittel & Eastman 1913, Lemaitre & McLaughlin 2009 MAK120528
The following text and image is copied verbatum from Wikipedia. Owing to limitations of time, the content has not yet been formatted or hyperlinked MAK120528

In evolutionary biology, carcinisation (or carcinization) is a hypothesised process whereby a crustacean evolves into a crab-like form from a non-crab-like form. The term was introduced by L. A. Borradaile, who described it as "one of the many attempts of Nature to evolve a crab".[1]
Carcinisation is believed to have occurred independently in at least five groups of decapod crustaceans, most notably king crabs, which most scientists believe evolved from hermit crab ancestors. The other examples are the family Porcellanidae, or porcelain crabs (these are closely related to squat lobsters), the hairy stone crab Lomis hirta, the coconut crab Birgus latro, and true crabs.[2] The example of king crabs (family Lithodidae) evolving from hermit crabs has been particularly well studied and, although some doubt this theory, there is considerable evidence in its favour. For example: most hermit crabs are asymmetrical, so that they fit well into spiral snail shells; the abdomens of king crabs, even though they do not use snail shells for shelter, are also asymmetrical.[3][4][5][6]
An exceptional form of carcinisation, termed "hypercarcinisation", is seen in the porcelain crab Allopetrolisthes spinifrons.[7] In addition to the shortened body form, A. spinifrons also shows similar sexual dimorphism to that seen in true crabs, where males have a reduced pleon compared to females.[7]
Image: The porcelain crab Neopetrolisthes resembles a crab, but is more closely related to squat lobsters and hermit crabs.
References
[1] Patsy A. McLaughlin & Rafael Lemaitre (1997). "Carcinization in the anomura – fact or fiction? I. Evidence from adult morphology". Contributions to Zoology 67 (2): 79–123. PDF
[2] C. L. Morrison, A. W. Harvey, S. Lavery, K. Tieu, Y. Huang & C. W. Cunningham (2001). "Mitochondrial gene rearrangements confirm the parallel evolution of the crab-like form". Proceedings of the Royal Society B: Biological Sciences 269 (1489): 345–350. doi:10.1098/rspb.2001.1886. PMC 1690904. PMID 11886621.
[3] C. W. Cunningham, N. W. Blackstone & L. W. Buss (1992). "Evolution of king crabs from hermit crab ancestors". Nature 355 (6360): 539–542. Bibcode 1992Natur.355..539C. doi:10.1038/355539a0. PMID 1741031.
[4] Patsy A. McLaughlin, Rafael Lemaitre & Christopher C. Tudge (2004). "Carcinization in the Anomura - fact or fiction. 2. Evidence from larval, megalopal and early juvenile morphology". Contributions to Zoology 73 (3): 165–205. hdl:7373
[5] Ling Ming Tsang, Tin-Yam Chan, Shane T. Ahyong & Ka Hou Chu (2011). "Hermit to king, or hermit to all: multiple transitions to crab-like forms from hermit crab ancestors". Systematic Biology 60 (5): 616–629. doi:10.1093/sysbio/syr063. PMID 21835822.
[6] Rafael Lemaitre & Patsy A. McLaughlin (2009). "Recent advances and conflicts in concepts of anomuran phylogeny (Crustacea: Malacostraca)" (PDF). Arthropod Systematics & Phylogeny 67 (2): 119–135.
[7] Alexandra Hiller, Carlos Antonio Viviana & Bernd Werding (2010). "Hypercarcinisation: an evolutionary novelty in the commensal porcellanid Allopetrolisthes spinifrons (Crustacea: Decapoda: Porcellanidae)" (PDF). Nauplius 18 (1): 95–102.
While there is now agreement that the Anomura, minus the Thalassinidea, constitute a monophyletic group, composition and relationships of the various lineages within this clade remain highly controversy. Several phylogenetic studies based on molecular data are particularily contradictory. (Lemaitre & McLaughlin 2009) implying either that the various groups evolved during a short period, but some time after the initial early Triassic radiation of the repantian decapods, or that the various taxa are insufficiently studied, artifacts haven't been corrected, and so on.
The following dendrogram is modified and updated (Lemaitre & McLaughlin 2009), from Christopher Taylor - Variety of LIfe. Not all groups are included MAK120530
References
[B01] Boyko, C. B. 2001. The identity of Hippa caerulea Risso 1816: An isopod in mole crab's disguise. Crustaceana 74 (1): 115-122.
[B02] Boyko, C. B. 2002. A worldwide revision of the recent and fossil sand crabs of the Albuneidae Stimpson and Blepharipodidae, new family (Crustacea: Decapoda: Anomura: Hippoidea). Bulletin of the American Museum of Natural History 272: 1-396.
[CA04] Casadío, S., A. de Angeli, R. M. Feldmann, A. Garassino, J. L. Hetler, A. Parras & C. E. Schweitzer. 2004. New decapod crustaceans (Thalassinidea, Galatheoidea, Brachyura) from the Middle Oligocene of Patagonia, Argentina. Annals of Carnegie Museum 73 (2): 25-47.
[H86] Holthuis, L. B. 1986. Decapoda. In Stygofauna Mundi: A Faunistic, Distributional, and Ecological Synthesis of the World Fauna inhabiting Subterranean Waters (including the Marine Interstitial) (L. Botosaneanu, ed.) pp. 589-615. E. J. Brill / Dr. W. Backhuys: Leiden.
[HC79] Huff, J. A., & S. P. Cobb. 1979. Penaeoid and sergestoid shrimps (Crustacea: Decapoda). Memoirs of the Hourglass Cruises 5 (4): 1-101.
[MD01] Martin, J. W., & G. E. Davis. 2001. An updated classification of the Recent Crustacea. Natural History Museum Los Angeles County, Science Series 39: 1-124.
[ML01] McLaughlin, P. A., & R. Lemaitre. 2001. A new family for a new genus and new species of hermit crab of the superfamily Paguroidea (Decapoda: Anomura) and its phylogenetic implications. Journal of Crustacean Biology 21 (4): 1062-1076..
Anomura MacLeay, 1838.
Synonym: Anomala De Haan, 1839
From the Jurassic
Phylogeny: Either (if (if Platykotta and Eocarcinus are basal or stem Anomura not related to crabs, in which case the present taxon becomes Crown Anomura)
Stem Anomura : Platykotta + (Eocarcinus + * : Hippoidea + (Aeglidae + Galatheoidea + Kiwa + Lomis + (Paguroidea + Lithodoidea))))
or (if Platykotta and Eocarcinus are basal Meiura rather than basal Anomura) Meiura : Platykotta + (Eocarcinus + (Brachyura + * : Hippoidea + (Aeglidae + Galatheoidea + Kiwa + Lomis + (Paguroidea + Lithodoidea)))
Characters:. Synapomorphies (from Dixon et al. 2003): Basal articles of second antenna fitting into a notch in the carapace; coxosternal joints of thoracopods reversed; cheliped rotated so that dactylus is horizontal; first pleopod of female absent. - CKT.
Comments: [1] from the Greek, anomoios unlike, different and oura, tail, referring to the fact taht these animals have a different tail (more properly abdomen) to the true crabs.
[2] In terms of priority, Anomura just pips Anomala at the post (1938 to 1839) (ref)
[3] Alternative phylogenies are used here owing to uncertainty over status of the two basal forms Platykotta and Eocarcinus. See Förster, 1985 and Krobicki & Zaton 2008 for the thesis that they are transitional between Triassic Pemphicidae and Jurassic Prosopid and Homolodromioid crabs (the most primitive crab lineages). See Feldmann & Schweitzer 2010 and Chablais et al 2011 for the thesis that they are basal Anomura and not in any way related to true crabs.
Thumbnail images for the following taxa are from the Wikipedia Anomura page MAK120530

Hippoidea Latreille, 1825
From the Late Cretaceous; ghost lineage suggests at least a Jurassic origin
Phylogeny: Anomura : (Aeglidae + Galatheoidea + Kiwa + Lomis + (Paguroidea + Lithodoidea)) + *)
Comments: sand crabs or mole crabs, remarkably convergent with raninid true crabs. In some recent phylogenetic analyses (Ahyong & O'Meally 2004; Porter et al 2005; Tsang et al. 2008) they are the most basal extant anomuran taxon
Image and link; Wikipedia

Galatheoidea Samouelle, 1819
From the Jurassic
Phylogeny: Anomura : Hippoidea + (Aeglidae + Kiwa + Lomis + (Paguroidea + Lithodoidea) + * )
Comments: The porcelain crabs, squat lobsters, and their relatives

No known fossils
Phylogeny: Anomura : Hippoidea + (Aeglidae + Galatheoidea + Lomis + (Paguroidea + Lithodoidea) + * )
Comments: a small but higher specialised group of marine decapods living at deep-sea hydrothermal vents and cold seeps. The animals are commonly referred to as "yeti lobsters" or "yeti crabs" (Wikipedia). Several species of the single genus are placed in the monotypal family Kiwaidae. This was originally placed in the Galatheoidea, but then elevated to its own superfamily rank (Kiwaoidea), although not all accepted this (Lemaitre & McLaughlin 2009)
From the Cretaceous; Recent restricted to of South America
Phylogeny: Anomura : Hippoidea + (Galatheoidea + Kiwa + Lomis + (Paguroidea + Lithodoidea) + * )
Comments: secondarily freshwater lobster-like forms currently restricted to South America, in the Cretaceous they may have had a Gondwana-wide distribution as fossils are known also from New Zealand and as far north as Mexico. Relations with other anomuran groups are uncertain; they have previously been included under the Galatheoidea but are now placed in their own superfamily. Several studies (e.g. Ahyong & O'Meally 2004, Porter et al 2005) place them as sister group to the monophyletic Lomisidae.
Lomis hirta (Lamarck, 1818)
Recent of southern Australia
Phylogeny: Anomura : Hippoidea + (Aeglidae + Galatheoidea + Kiwa + (Paguroidea + Lithodoidea) + * )
Comments: The hairy stone crab, Lomis hirta (monotypal family Lomisidae and superfamily Lomisoidea Bouvier, 1895; the name Lomidae is incorrect), is a slow-moving crab-like animal covered in brown hair. Its relations to other anomuran groups are unclear, but represents a separate case of carcinisation - Wikipedia.

Paguroidea Latreille, 1802
From the Jurassic
Phylogeny: Anomura : Hippoidea + (Aeglidae + Galatheoidea + Kiwa + Lomis + (Lithodoidea + * )
Comments: The hermit crabs, a familiar group of decapod crustaceans, with over a thousand extant species. They occupy empty gastropod shells which protect the sSoft, coiled, asymmetrical abdomen. Mesozoic forms seem to have preferred ammonites, which have a planospiral shell, and so they would have had symmetrical abdomens.

From the Jurassic
Phylogeny: Anomura : Hippoidea + (Aeglidae + Galatheoidea + Kiwa + Lomis + (Paguroidea + * ))
Comments: King crabs, somewhat spider-crab-like anomurans thought to have evolved from hermit crabs, and previously included with them, but now placed in a separate superfamily, Lithodoidea
page MAK120529