A is happy. B is not.
Yet B is worthy, I dare say
Of more prosperity than A . . .
But should A perish?
That should he --
That is, assuming I am B
 .
.| Cretaceous | ||
| Mesozoic Era | The Cretaceous - 2 |
| Jurassic | Paleozoic | Mesozoic | ||
| Paleogene | Cenozoic | Early Cretaceous I | Timescale |
| Mesozoic Triassic Jurassic Cretaceous Early Cretaceous I (Neocomian) Early Cretaceous II (Aptian-Albian) Late Cretaceous I High Cretaceous) Late Cretaceous II (End-Cretaceous) Cenozoic Paleogene |
The Cretaceous |
See how the fates their gifts alot:
A is happy. B is not.
Yet B is worthy, I dare say
Of more prosperity than A . . .
But should A perish?
That should he --
That is, assuming I am B.
-- William S. Gilbert, The Mikado
Aberrant bivalve mollusks, known as rudists, dominated Cretaceous reefs. You can find more information, as well as clumsy evasions and outright speculation, in the sections on the Neocomian and the Aptian. Here, after some perfunctory remarks, we're going to take up the fate of another legion of reef builders, the scleractinian corals. We focus on corals, not only because they are important today, but also to investigate the role of ecological competition in evolution.
Here's the problem. In the old days -- perhaps before 1980 or so -- most scientists believed that evolution was driven largely by competitive exclusion: the "nature red in tooth and claw" school of evolutionary theory. Taxon B replaced A because it was bigger, badder, and more "fit." Gradually, the pendulum swung the other way, as evidence accumulated suggesting that blind luck, climate, and environmental factors played an enormous role. Today, orthodoxy requires every paper to conclude that B replaced A only because A was already going to hell in a hand basket for environmental reasons. It is now considered very bad taste to contemplate that B might have been better at anything, or more "fit," than A, or even to admit that A and B actually competed for anything.
 It may be simple perversity, or the fact that we are bad-tempered and easily irritated, but it is striking that these two views of the engines of evolution just happen to parallel the educational orthodoxy of academia, as it has changed over the years. Our suspicions are aroused because so much of this discussion seems to be over semantics, rather than science. Consider some examples. If I say that B is more fit than A, might I mean that, by chance, the environment changed in a way that promoted B's survival over A? Alternatively, if I relate that A was already near-extinct before B moved in, didn't B still have to out-compete C, D, and E for the open slot? As always, we meticulously avoid taking any position on the underlying philosophical positions. The problem here is that both philosophical positions seem to be getting in the way of a more dispassionate assessment of the data.
It may be simple perversity, or the fact that we are bad-tempered and easily irritated, but it is striking that these two views of the engines of evolution just happen to parallel the educational orthodoxy of academia, as it has changed over the years. Our suspicions are aroused because so much of this discussion seems to be over semantics, rather than science. Consider some examples. If I say that B is more fit than A, might I mean that, by chance, the environment changed in a way that promoted B's survival over A? Alternatively, if I relate that A was already near-extinct before B moved in, didn't B still have to out-compete C, D, and E for the open slot? As always, we meticulously avoid taking any position on the underlying philosophical positions. The problem here is that both philosophical positions seem to be getting in the way of a more dispassionate assessment of the data.
It turns out that Cretaceous corals are an interesting study for these purposes for two reasons. First and foremost, some very good studies have been done over the last few years by some rather careful workers. It's nice to have good data. Second, there's no dispute that corals were rather marginal reef-builders in the later Cretaceous and that rudist bivalves occupied the center stage. What can we say about competition and co-existence in this system?
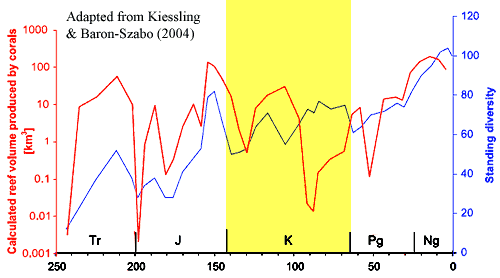
We focus first on Kiessling & Baron-Szabo 2004). These authors were mainly interested in re-examining some quantitative conclusions reached by Sepkoski relating to the effect of the KT extinction on corals. In the process, they pull together the data shown in the figure, largely from previous work [1]. This graph charts coral reef formation rates (productivity) and coral diversity over the course of the Mesozoic. Note that the scale for reef productivity rate is logarithmic, so that the strong peaks and valleys in the reef productivity graph are even more dramatic than they appear at first glance. For the Cretaceous, coral diversity showed a slight, but relatively steady increase. Coral reef productivity was low throughout the Cretaceous; but it was essentially zero during the Late Cretaceous, except at the very end of the epoch. For our purposes, the more interesting pattern is the correlation between diversity and productivity. For all epochs except the Late Cretaceous and Paleocene, productivity and diversity are closely correlated. For the 50 My comprising the Late Cretaceous and Paleocene, the relationship is reversed. What's going on? Rather than answer our rhetorical question, we will deepen the mystery, or at least the confusion, by producing another messy, compound graph:
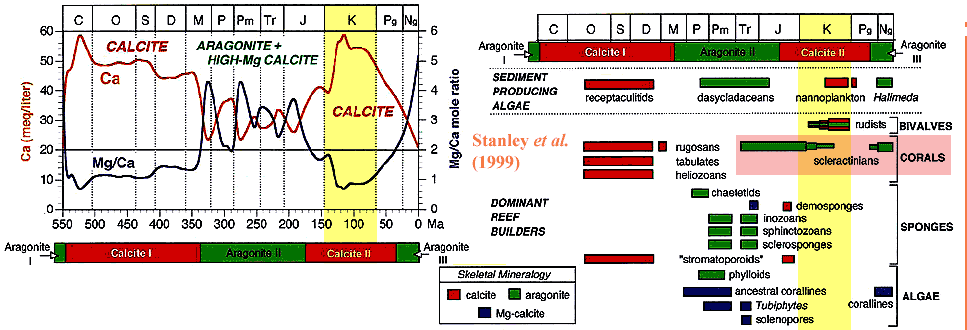
One of the most successful efforts to explain the ups and downs of reef-building taxa relies on marine calcium- magnesium balance. These two divalent cations (Ca++ and Mg++) are chemically similar and may substitute for each other in crystal structures. This substitution has some biological importance, since the calcium atoms in the calcium carbonate (CaCO3) of shells may be partially substituted by magnesium. This mixed material is called high-magnesium calcite. Perhaps more importantly, calcium carbonate can crystallize in one of two ways, calcite and aragonite. When magnesium levels are high, aragonite is the favored crystal form. Aragonite is less stable than calcite. However, organisms that are able to use aragonite have an advantage when magnesium levels are high. This is shown in the graph above, adapted from Stanley et al. (1999). The left half shows calcium levels and the magnesium/calcium ratio aver the entire Phanerozoic. The right half shows the dominant reef builders over the same scale. In broad, the correspondence between ion balance and reef-builder communities is remarkable. Reef-builders seem to be adapted to use either calcite or aragonite in their structural framework. When the calcium- magnesium ratio changes, so do the reef-builders.
 But perhaps a closer look is warranted. The "aragonitic" scleractinian corals seem to have done remarkably well in the Early Cretaceous, all things considered, particularly in view of the fact that calcium levels were at an all-time high. Stanley et al. show the scleractinian corals petering out over the Early Cretaceous, presumably in response to changing ion balance. But we have a dark and suspicious nature. Out of curiosity, we combined the graphs of Kiessling & Baron-Szabo together with the data from Stanley et al. for the Mesozoic.
But perhaps a closer look is warranted. The "aragonitic" scleractinian corals seem to have done remarkably well in the Early Cretaceous, all things considered, particularly in view of the fact that calcium levels were at an all-time high. Stanley et al. show the scleractinian corals petering out over the Early Cretaceous, presumably in response to changing ion balance. But we have a dark and suspicious nature. Out of curiosity, we combined the graphs of Kiessling & Baron-Szabo together with the data from Stanley et al. for the Mesozoic.
The results are quite peculiar. From the beginning of the Mesozoic, right through the Early Cretaceous, all of our metrics are closely correlated: coral diversity, coral biomass, and calcium concentration -- almost precisely the opposite of the conclusions of Stanley et al. That is, the graph shows that early Mesozoic corals were not happy with magnesium. To the contrary, coral diversity and biomass follow calcium levels very closely. As with the diversity/biomass curves, this correlation falls apart in the Late Cretaceous. Then, after the Paleocene, the correlation reverses. Diversity and biomass follow each other, with both rising as calcium levels fall.
How do we explain all this? Should we, perhaps, have stuck to showing pretty pictures of ammonites and blown off the temptation to look at the actual data? Too late for that now, I suppose.
But the case is not hopeless. First, we can tease apart the biomass and diversity curves. Corals occur as solitary and colonial forms. It is the colonial forms -- or some of them -- that contribute to global reef biomass. The data from Kiessling & Baron-Szabo show clearly that the Cretaceous and Paleogene were actually moderately happy times for scleractinian corals, with slow but steady increases in diversity. But, what the data also indicate is that the Late Cretaceous and Late Paleocene were really rotten times for a particular ecological (not phylogenetic) group of corals, the reef builders.
The simplest explanation for these data is competitive exclusion by rudists. Rudist diversity really escalated beginning in Aptian-Albian time and ended, abruptly, at the KT. The rudists didn't devour scleractinians, or hold any personal grudges. They just took up room in an increasing number of reef ecospaces. It shouldn't be a surprise that solitary corals continued to thrive as before. This happened precisely because the engine was competitive exclusion of a particular ecotype, not an environmental stressor applicable to corals generally.
With that background, the calcium curve makes more sense. We need only abandon the untenable belief that Early Mesozoic corals were magnesium lovers. The data are simply to the contrary.Scleractinian diversity and biomass follow the calcium curve from the first appearance of scleractinians, at the beginning of the Middle Triassic, right through the Neocomian. However, the big calcium spike of Aptian-Albian time did not result in a corresponding spike in coral biomass because rudist competition was already beginning to take hold, suppressing the recovery. The complex pattern around the KT is probably a combination of various effects, which cannot easily be untangled at this level of temporal resolution. These effects would include:
a) The initial diversification of several new families of truly aragonite corals, such as the Flabellidae, Meandrinidae, and Dendrophyllidae Stanley 2003);
b) The KT extinction itself;
c) The removal of rudist competition across the KT;
d) The gradual reduction in calcium concentrations over the End-Cretaceous; and
e) The resulting replacement of Mesozoic calcite genera by Cenozoic aragonitic genera.
The bottom line seems to be that, to the extent one can separate environmental and competitive effects in this system, both are important. However, competition and environmental effects may be limiting in different ways and at different times. It is interesting that coral and rudist communities managed to coexist for about 40 My in the Early Cretaceous, presumably by ever-finer partitioning of the available ecospace. However, at some point, competitive exclusion became the dominant mode. These data suggest that coral families adapted to prefer aragonite did not evolve until the end of the Mesozoic. Prior to that time, scleractinian corals preferred high calcium levels. So, Cretaceous ion balances cannot explain their retreat in the face of rudist competition.
Images: Rudist reef cross-section from the M08313 Sedimentology site of Oxford Brookes University.
ATW050216. Revised ATW050516. Text public domain. No rights reserved.
plants
Although the Jurassic floras of ferns, cycads, and conifers continued, the most dramatic occurrence during the Cretaceous, was the first appearance of the flowering plants, the angiosperms. By the end of the Cretaceous, a number of modern plant forms had evolved.
 marine invertebrates
marine invertebrates
 Ammonites, belemnites left), other molluscs were abundant, as were sponges, bivalves and echinoids. Several new groups of Echinoids appear during this time, and the families Echinocorythidae and Spatangidae become abundant. The brachiopods meanwhile declined in diversity. Higher crustaceans such as lobsters become common. Corals were of essentially modern types. Modern gastropod groups (especially Neogastropods) appear. These differ from all previous gastropods by being largely carnivorous, and, like the Mesogastropods, are easily able to live in sandy environments.
Ammonites, belemnites left), other molluscs were abundant, as were sponges, bivalves and echinoids. Several new groups of Echinoids appear during this time, and the families Echinocorythidae and Spatangidae become abundant. The brachiopods meanwhile declined in diversity. Higher crustaceans such as lobsters become common. Corals were of essentially modern types. Modern gastropod groups (especially Neogastropods) appear. These differ from all previous gastropods by being largely carnivorous, and, like the Mesogastropods, are easily able to live in sandy environments.
The normally conservatively coiled ammonoids take on bizarre shapes, which was at one time attributed to a sort of racial senility. This idea is no longer considered valid. In addition to uncoiled ammonoids, and normal coiled forms, both with complex suture patterns, there are those types found in areas of North Africa, Syria, South America and southern USA in which the suture-lines had become simplified so that they closely resembled those of the Triassic Cerratites. These are, naturally, called "pseudo-ceratites".
Cretaceous reefs are largely made up of rudists, huge bivalves with one cone-shaped valve and the other reduced to a small lid-like structure. A single animal could be a meter in height. Rudists grew rapidly, most certainly with the help of symbiotic algae. The early Cretaceous forms like Monopleura and Caprotina were replaced during the later Cretaceous by Hippurites, whose growth-habit paralleled that of Rugose corals. Especially in the Mediterranean region, these were an important part of the marine ecosystem.
fish
Modern teleost fishes become widespread around the middle and especially towards the end of the period, and modern sharks also appear during this time.
terrestrial invertebrates
The appearance of flowering plants greatly stimulated insect evolution, as the plants used insects for pollination. Many modern groups of insects appeared and began to diversify, including the oldest known ants and butterflies.
Amphibia:
![]() Apart from a single lineage of giant temnospondyls confined to the rift valleys of south-east Gondwana, the amphibians are represented only by existing groups such as frogs and salamanders
Apart from a single lineage of giant temnospondyls confined to the rift valleys of south-east Gondwana, the amphibians are represented only by existing groups such as frogs and salamanders
Reptilia (non-archosauria)
The chelonia turtles and tortoises) are common, and include representatives of both living and extinct groups. Some marine forms attained enormous size, such as the 3 meter Archelon.
 The tuatara-like Sphenodontia are much reduced, their ecological role being taken by the Squamata (lizards and snakes). One line of aquatic lizards evolve into gigantic marine forms, the predatory Mosasauridae, with lengths of up to 10 meters and more.
The tuatara-like Sphenodontia are much reduced, their ecological role being taken by the Squamata (lizards and snakes). One line of aquatic lizards evolve into gigantic marine forms, the predatory Mosasauridae, with lengths of up to 10 meters and more.
The Ichthyosauria are greatly reduced, with only a single family straggling through to the early part of the late Cretaceous. It is likely that they were unable to adapt to the new fast-swimming teleost fish. They died out long before the Cretaceous ended.
 Plesiosaurs are represented by a number of new forms, including the Polycotylidae, Cimoliasauridae, and huge long-necked Elasmosaurids left).
Plesiosaurs are represented by a number of new forms, including the Polycotylidae, Cimoliasauridae, and huge long-necked Elasmosaurids left).
Crocodiles
Modern crocodilians Eusuchia) evolve for the first time, while a number of more primitive Mesouchian groups continue. The sea crocodiles die out early in the Cretaceous. The warm tropic conditions however are good news for the fresh-water crocs, some of which (e.g. Sarcosuchus, Deinosuchus) attain huge size. These gigantic crocodiles doubtless fed on unwary dinosaurs in the same way that large modern crocodiles will grab an ungulate mammal (such as a zebra or wildebeest) that approaches the water to drink.
Dinosaurs and birds
 Like the Jurassic, the Cretaceous Period is distinguished in the popular imagination by the presence of great dinosaurs, marine reptiles, and flying pterosaurs. Many new types of dinosaurs evolved at this time. Among the greatest of the giants, the brachiosaurs and diplodocids died out, while new sauropod dinosaurs, specifically the Titanosauria, continued to flourish in Gondwanaland, but became rare in Laurasia. The plated Stegosauria were replaced by the heavily armoured Ankylosauria. The Ceratopsia (horned dinosaurs) evolved and flourished in Asiamerica but (apart from one or two questionable forms) are unknown elsewhere. The Pachycepalosauria seem to have been similarly limited in distribution. Curiously, the most abundant and diverse of the Cretaceous herbivores were the unarmoured ornithopod dinosaurs, specifically the hypsilophodontids and iguanodontian lines, all of which achieved cosmopolitan distribution.
Like the Jurassic, the Cretaceous Period is distinguished in the popular imagination by the presence of great dinosaurs, marine reptiles, and flying pterosaurs. Many new types of dinosaurs evolved at this time. Among the greatest of the giants, the brachiosaurs and diplodocids died out, while new sauropod dinosaurs, specifically the Titanosauria, continued to flourish in Gondwanaland, but became rare in Laurasia. The plated Stegosauria were replaced by the heavily armoured Ankylosauria. The Ceratopsia (horned dinosaurs) evolved and flourished in Asiamerica but (apart from one or two questionable forms) are unknown elsewhere. The Pachycepalosauria seem to have been similarly limited in distribution. Curiously, the most abundant and diverse of the Cretaceous herbivores were the unarmoured ornithopod dinosaurs, specifically the hypsilophodontids and iguanodontian lines, all of which achieved cosmopolitan distribution.
 Among the theropod carnivores there was an astonishing radiation of large and small bird-like forms, ranging from the giant carcharodontosaurs, spinosaurs, and Tyrannosaurs, through the medium-sized sickle-clawed deinonychids to the bird-like ornithomimosaurs and the enigmatic segnosaurs. Alongside these medium to large forms were an extraordinary range of birds and dinosaur-like protobirds. Indeed, during the Cretaceous it would have been impossible to tell where dinosaurs ended and birds began, such is the continuum between them.
Among the theropod carnivores there was an astonishing radiation of large and small bird-like forms, ranging from the giant carcharodontosaurs, spinosaurs, and Tyrannosaurs, through the medium-sized sickle-clawed deinonychids to the bird-like ornithomimosaurs and the enigmatic segnosaurs. Alongside these medium to large forms were an extraordinary range of birds and dinosaur-like protobirds. Indeed, during the Cretaceous it would have been impossible to tell where dinosaurs ended and birds began, such is the continuum between them.
 The birds seem to have pushed the Pterosaurs flying reptiles) progressively to extinction. As the Cretaceous progressed the Pterosaurs declined steadily, until at the end of the period only one or two aberrant giant forms (Pteranodon, Quetzelcoatlus) remained.
The birds seem to have pushed the Pterosaurs flying reptiles) progressively to extinction. As the Cretaceous progressed the Pterosaurs declined steadily, until at the end of the period only one or two aberrant giant forms (Pteranodon, Quetzelcoatlus) remained.
Many new mammal groups also appeared during this time, including the three living groups of mammals placentals, marsupials and monotremes) which appear during the mid Cretaceous.
The Cretaceous-Tertiary Extinction
The Cretaceous and the entire Mesozoic was brought to an end by one of the greatest mass extinctions of all time, the K-T event or terminal Mesozoic extinction. About one half of all animal families died out, including dinosaurs, many marine reptiles, several lines of archaic birds, the ammonoid and most belemnite cephalopods, rudist clams, and many microorganisms.
University of California Museum of Paleontology -- The Cretaceous Period
Cretaceous map(50.9 kb)
Geophysicists model the Cretaceous motions of Australia
More Links: Enchanted Learning and Wikipedia have good, very accessible pages on the Cretaceous. Cretaceous Tethyan Stratigraphy has detailed stratigraphic information and some maps. The USGS covers North American stratigraphy, at a less technical level. All things Cretaceous has very well-organized links and some connecting material. We find the tone of the site irritatingly slick -- but, then, we are very easily irritated. ATW060107