 Cercozoa, originally named by Cavalier-Smith in 1998, is a diverse group of taxa united solely on molecular grounds, but supported by a number of genes (Longet et al., 2003). As generally circumscribed, Cercozoa also includes Gromiidae and Phytomyxea, but these more divergent taxa have been listed separately here to show their relative phylogenetic positions (and also to avoid having to lump Foraminifera in with the Cercozoa). For a brief period before 1998, the clade soon to be called Cercozoa was referred to as Rhizopoda, as it included a large proportion of the species previously included in that form-taxon (specifically those bearing filose pseudopodia). But as many rhizopods were not in this group, including the best-known example, Amoeba, and many Cercozoa are flagellates rather than amoeboid, the name Cercozoa is much more welcome. Cercozoa, originally named by Cavalier-Smith in 1998, is a diverse group of taxa united solely on molecular grounds, but supported by a number of genes (Longet et al., 2003). As generally circumscribed, Cercozoa also includes Gromiidae and Phytomyxea, but these more divergent taxa have been listed separately here to show their relative phylogenetic positions (and also to avoid having to lump Foraminifera in with the Cercozoa). For a brief period before 1998, the clade soon to be called Cercozoa was referred to as Rhizopoda, as it included a large proportion of the species previously included in that form-taxon (specifically those bearing filose pseudopodia). But as many rhizopods were not in this group, including the best-known example, Amoeba, and many Cercozoa are flagellates rather than amoeboid, the name Cercozoa is much more welcome.
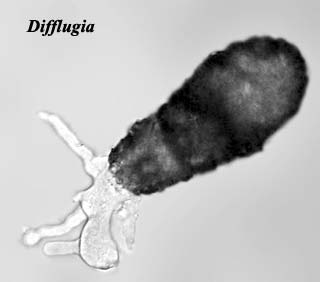
Amongst notable members of the Cercozoa are amoeboid forms such as Difflugia, which produce agglutinated tests that may be fossilised (the record extends back to the Neoproterozoic
– Finlay et al., 2004), and the Chlorarachnea (e.g. Chlorarachnion), marine amoeboid organisms which possess chloroplasts derived from a secondary endosymbiosis with a green alga. Cavalier-Smith, 2003). The nucleus of the endosymbiont in Chlorarachnion, in fact, has not fully degraded as in most secondarily plastid-bearing eukaryotes, and the chloroplast retains a small nucleomorph contained within the surrounding membranes.
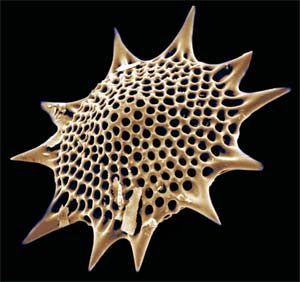
Nikolaev et al. (2004) and Polet et al. (2004) both found Phaeodarea to also be nested within Cercozoa, though a strong association with any particular taxon or taxa was not supported. Phaeodarea were traditionally included in Radiolaria, and share with Acantharea and Polycystinea the traits of a glassy shell (formed of a combination of silica and organic material in Phaeodarea) and a capsule dividing the cytoplasm into inner and outer compartments. In the Radiolaria as here defined, however, the capsule is thin and perforated by numerous pores – in Phaeodarea, the capsule is much thicker, and usually only three pores pass through it, the astropylum and and usually two parapyla situated at the opposite pole. The astropylum forms a cone-like cytopharynx that is used for the ingestion of food items. Phaeodarea also bear a phaeodium, consisting of balls of darkly pigmented waste matter, usually near the astropylum. Phaeodarea also lack algal endosymbionts and cross-branches between the axopods. Polet et al. (2004).
Image: Difflugia from the Droplet site.
Text © Christopher Taylor 2004. CT041218 |