The Fungi
What are the Fungi?
 The Fungi are the great saprophytes, the master recyclers. They are the black rot, the dry rot, and the white rot, the colorful fate of last week's lasagna left too long in the 'fridge, and the great, grey walls of stinking mould that can destroy whole buildings. But, they are also the baker's yeast and the brewer's yeast. They are the difference between grape juice and Chateauneuf du Pape. They are the portobellos and the morels and the cloud ears and the truffles. In fact, the French could not be half so obnoxious about their cuisine were it not for the Fungi. But, then again, perhaps they could [1].
The Fungi are the great saprophytes, the master recyclers. They are the black rot, the dry rot, and the white rot, the colorful fate of last week's lasagna left too long in the 'fridge, and the great, grey walls of stinking mould that can destroy whole buildings. But, they are also the baker's yeast and the brewer's yeast. They are the difference between grape juice and Chateauneuf du Pape. They are the portobellos and the morels and the cloud ears and the truffles. In fact, the French could not be half so obnoxious about their cuisine were it not for the Fungi. But, then again, perhaps they could [1].
We leave that conundrum for another day. The first order of business ought to be the matter of definition. How do we define this group? We have found no hint that anyone is using a workable phylogenetic definition of the Fungi. A phylogenetic definition, for those who have somehow managed to escape our interminable, high-pitched whining on the subject, is a definition based on some explicit hypothesis about a group's relative position in phylospace. For example, dinosauria is defined as the last common ancestor of Triceratops and birds and all descendants of that ancestor. This may be conveniently abbreviated: "Triceratops + birds". Such a definition is quite different from a definition based on some arbitrary set of characteristics which approximate an implicit, unstated, and therefore untestable notion of what a dinosaur "ought" to look like. That second type of definition is referred to as an "apomorphy-based" definition. It is properly viewed with the same derisive contempt with which M. Auguste Escoffier (at right) would regard the use of corn starch to thicken a demi-glace [3].
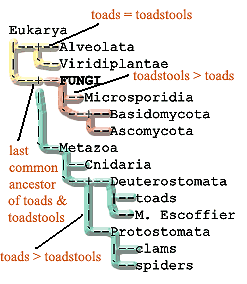 Were we in a position to impose a phylogenetic definition on the Fungi, our leading candidate would be the stem group "toadstools > toads" (all organisms more closely related to Basidiomycota than to Tetrapoda [2]). That definition presupposes a close relationship between Metazoa and Fungi. However, such an assumption shouldn't slow us up much. The Metazoa-Fungi connection now seems quite secure. This definition would, however, require us to gather the Microsporidia into the brotherhood of the Fungi. Microsporidia could be Fungi under many definition of the taxon, but they are certainly closer to Fungi than to toads. Such a definition would also dispense with meaningless arguments about the inclusion of the Chytridiomycota within Fungi.
Were we in a position to impose a phylogenetic definition on the Fungi, our leading candidate would be the stem group "toadstools > toads" (all organisms more closely related to Basidiomycota than to Tetrapoda [2]). That definition presupposes a close relationship between Metazoa and Fungi. However, such an assumption shouldn't slow us up much. The Metazoa-Fungi connection now seems quite secure. This definition would, however, require us to gather the Microsporidia into the brotherhood of the Fungi. Microsporidia could be Fungi under many definition of the taxon, but they are certainly closer to Fungi than to toads. Such a definition would also dispense with meaningless arguments about the inclusion of the Chytridiomycota within Fungi.
To our discredit, the foregoing discussion may serve as a useful study in the use of the English conditional mode for advanced students of the language, but it ignores the realities of fungal phylogeny. That reality is illustrated in the following examples.
 The "fungi contain cell walls and produce spores." Madigan et al. (2003). So ferns are Fungi?
The "fungi contain cell walls and produce spores." Madigan et al. (2003). So ferns are Fungi?
The Tree of Life will not venture even this far. It contents itself with a list of common names: "The organisms of the fungal lineage include mushrooms, rusts, smuts, puffballs, truffles, morels, molds, and yeasts, as well as many less well-known organisms." In other words, the fungi are defined by listing a number of vague, vernacular terms with a completely indefinite catch-all category at the end.
"What is Fungi? Fungi are a group of organisms and micro-organisms that are classified within their own kingdom, the fungal kingdom, as they are neither plant nor animal. Fungi draw their nutrition from decaying organic matter, living plants and even animals. They do not photosynthesize as they totally lack the green pigment chlorophyll, present in green plants. Many play an important role in the natural cycle as decomposers and return nutrients to the soil, they are not all destructive." WHAT IS Fungi? But this description would apply equally as well to most bacteria.
 "These nonmotile eukaryotes lack flagella and develop from spores." Dr. Fungus- Fungi, Fungus, Fungal. But see the image.
"These nonmotile eukaryotes lack flagella and develop from spores." Dr. Fungus- Fungi, Fungus, Fungal. But see the image.
We could beat this drum for quite a long time. The point is that, of the hundreds of references and sites on the web which purport to discuss the Fungi, not one of the many we have reviewed supplies a reasonable definition. Some sources are very useful in listing numerous characteristics of Fungi. But, the more characters listed, the more Fungi (in any phylogenetic sense) they exclude. A substantial majority of sources simply dodge the issue.
Ultimately, we are left in the untenable position of admitting that there is no definition in general use for the word "fungus." Happily, this yawning gap at the threshold of mycology seems to bother mycologists even less than it bothers the Fungi themselves. Thus, in a manner sanctioned by the universal practice of man and mushroom alike, we will pointedly ignore the yawning abyss at our feet, and move on to other matters.
ATW041112. Text public domain. No rights reserved.
We never eat bread cookies
For cookies have yeast,
And one little bite
Turns a man to a beast
O, can you imagine
A sadder disgrace
Than a man in the gutter
With crumbs on his face?
-- Song of the Salvation Army (trad.)
So, what about all those characteristics mentioned in the last section? The following is a list of the most commonly cited characters shared by most Fungi:
- The Fungi are eukayotes, which may exist in nature as either single and multi-celled organisms, or in both at different points in the the life cycle.
- Fungi are avascular -- no specialized respiratory, digestive or transport systems beyond the hyphae themselves.
- Most fungi grow as tubular filaments called hyphae. A connected mass of hyphae is a mycelium.
- Fungi have a vegetative body called a thallus, composed of hyphae.
- The walls of hyphae are often reinforced with chitin, a polymer of N-acetylglucosamine.
- Fungal cell membranes contain ergosterol, rather than cholesterol.
- The Fungi have a unique biosynthetic pathway for lysine.
- Fungi produce a unique form of tubulin in connection with nuclear division.
- Fungi have small nuclei with very little repetitive DNA
- Mitosis occcurs without dissolution of the nuclear membrane.
- Fungi are never autotrophs. No fungus has chlorophyll or chloroplasts.
- Fungi are usually found either as opportunistic saprophytes living on dead organic matter) or in some parasitic or symbiotic relationship with plants or other autotroph.
- Fungi digest food outside their bodies: they release enzymes into the surrounding environment (exoenzymes), breaking down organic matter into a form the fungus can absorb
- food reserves stores as glycogen (like animals), not starch (like plants).
- Fungi reproduce by means of spores, budding, or fragmentation.
- Spores may be either sexual or aesexual.
- Spores may be used as a dormant, resting phase, like bacterial spores
In short, Fungi are a rather odd, and distinctly different, part of the tree of life.
The following is our usual diversity table, which somewhat overemphasizes the basal Fungi. Recent work suggests that fungal diversity may be undersampled even at the highest taxonomic levels. Specifically, a taxonomic survey of alpine fungal communities which flourished under snow cover suggests that there may be 1-2 high-level fungal taxa between Basidiomycota and Ascomycota. Schadt et al. (2003).
Fungi |
| |
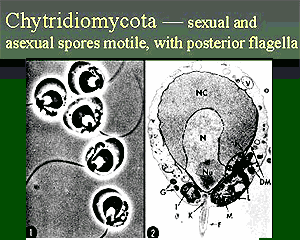
 The chytridomycotes, or "chytrids," are usually aquatic, either marine or freshwater. Presumably this is the original domain of the chytrids, and of all Fungi, but chytrids are also found in terrestrial communities almost as soon as there were terrestrial communities to be found in. So, for example, several different groups of chytrids are known from the Early Devonian Rhynie Chert. The implication is that they had begun to radiate even before the Devonian. They are a remarkably diverse lot, as one might expect from a basal radiation of the Fungi, and there is some possibility that the Chytridomycota may be paraphyletic, i.e. that all Fungi are descended from chytrids. The chytridomycotes, or "chytrids," are usually aquatic, either marine or freshwater. Presumably this is the original domain of the chytrids, and of all Fungi, but chytrids are also found in terrestrial communities almost as soon as there were terrestrial communities to be found in. So, for example, several different groups of chytrids are known from the Early Devonian Rhynie Chert. The implication is that they had begun to radiate even before the Devonian. They are a remarkably diverse lot, as one might expect from a basal radiation of the Fungi, and there is some possibility that the Chytridomycota may be paraphyletic, i.e. that all Fungi are descended from chytrids.
The chytrids are mostly single-celled forms, traditionally classified as protists. In fact, some sources still classify them with the Chromista, even though, so far as we can tell, chytrids have no light-sensitive pigments at all. What chytrids do have is a single- celled zoospore with an anterior flagellum, which is distinctly odd for a fungus. In fact, chytrids are the only large taxon of Fungi which produces a zoospore of any kind.
However, there's no real doubt about their position any more. For example Borneman & Hartin (2000) showed that rDNA primers from all of four basic fungal phyla (Trichomycota was not included) permit amplification of rDNA in the other fungal groups, including Chytridomycota, but don't amplify anything else. This strongly suggests that that rDNA from all four groups was very similar and that all are closely related.
That same conclusion can be reached for any number of other reasons. Chytrids have an absorptive mode of nutrition, like other Fungi. Chytrids have cell walls composed of chitin. Chytrids form hyphae. They share, with the other Fungi, key enzymes and metabolic pathways that are not found in other fungus-like protozoans (slime molds and water molds), in addition to oddities of molecular structure. Alkemar & Nygard (2003). The chytrids are surely the most basal Fungi, but Fungi they are.
Image Credit: The Microbial World.
ATW041115. Text public domain. No rights reserved. |
|
Unnamed Clade
| This group is sometimes referred to as Zygomycota, with Zygomycetes and Trichomycetes treated as included taxa. |
|
 Zygomycotes, like chytrids, are known from the Rhynie Chert, although, in this case, the identification is more tentative. What are actually seen are fungal hyphae which appear to pierce other cells, a characteristic of many zygomycotes. Definitive zygomycotes are found in Carboniferous exposures. A more familiar present-day example of a zygomycote is Rhizopus, the black bread mold. Zygomycotes, like chytrids, are known from the Rhynie Chert, although, in this case, the identification is more tentative. What are actually seen are fungal hyphae which appear to pierce other cells, a characteristic of many zygomycotes. Definitive zygomycotes are found in Carboniferous exposures. A more familiar present-day example of a zygomycote is Rhizopus, the black bread mold.
The Zygomycota are named for their characteristic teleomorph, which is referred to as a zygosporangium. The images at the glossary entry for gametangium are of Phycomyces and Rhizopus, both zygomycotes. They illustrate how the zygosporangium is formed from the head-on meeting of two hyphae whose ends have specialized as gametangia. The contents of the gametangia are mixed in the zygosporangium, which develops between them. The haploid nuclei from the gametangia then fuse. The zygosporangium develops a hard, thick chitin shell, which is frequently ornamented and may bear spines or other appendages. The remains of the gametangia protrude from the sides and are referred to as suspensors. The zygosporangium also serves as a resting phase, which will develop when conditions are favorable.
Zygomycotes also reproduce asexually. The haploid spores develop in a bulbous mitosporangium at the tips of specialized vertical hyphae referred to as sporangiophores.
Image: Dr. Paul Davis, Univ. N. Alabama.
ATW041116. Text public domain. No rights reserved. |
|
| From time to time we find a web resource that is so comprehensive well-organized that there is no point in providing a summary here. That is the case with the Trichomycota. We happily defer to The Trichomycetes: Fungal Associates of Arthropods. The authors' own summary of their work is Chapter 1 of the treatise. Trichomycotes are obligate commensals sometimes parasites) of insects and are thought to have developed with neopteran insects in the Mississippian. |
|
 The basidiomycotes are the rusts, smuts, gilled mushrooms, puffballs, stinkhorns, and club, shelf or coral fungi. They are one of the two major divisions of Fungi, the other being the Ascomycota. Definitive Basidiomycote fossils are known from the Late Devonian, although there has been a recent report of a possible Early Devonian lichen incorporating a probable basidiomycote fungus. The basidiomycotes are the rusts, smuts, gilled mushrooms, puffballs, stinkhorns, and club, shelf or coral fungi. They are one of the two major divisions of Fungi, the other being the Ascomycota. Definitive Basidiomycote fossils are known from the Late Devonian, although there has been a recent report of a possible Early Devonian lichen incorporating a probable basidiomycote fungus.
The Basidiomycota is such a large and diverse group, that the living members have little in common. The basidiomycote life cycle has a four unique properties which are probably synapomorphies, but which are no longer shared by all members of the group:
(1) The taxon is named for the basidium where sexual spores are produced.
(2) The life cycle generally includes a persistent dikaryon, frequntly large (e.g., a mushroom) in which each cell in the thallus contains two haploid nuclei, typically as the resulting of a mating event
(3) Clamp connections (explained at the glossary entry) are unique to Basidiomycota and are used to maintain the dikaryon state during hyphal division.
(4) Many basidiomycotes can launch spores into the air in a process referred to as ballistospory.
The basidiospores bear a single haploid nucleus. They germinate into hyphae with a single nucleus in each compartment, a monokaryon. A mating event results from end-to-end fusion of hyphae, as in Zygomycota, or fusion of a hypha with an oidium, a specialized mating spore. Then the resulting dikaryon divides through clamp connections so that the dikaryon state is maintained. Many basidiomycotes remain in the dikaryon state indefinitely. Under appropriate conditions, the dikaryon will produce fruiting bodies. Some of these hyphae produce basidia, such as the cells lining the "gills" under the cap of gilled mushrooms. Ultimately, the two haploid nuclei in each basidium fuse (karyogamy) to form a diploid nucleus. This then undergoes meiosis to produce four haploid nuclei whch migrate into the basidiospores and are dispersed into the environment.
ATW041116. Text public domain. No rights reserved. |
|
 The Ascomycota are the largest and most diverse group of Fungi. They include the yeasts, most of the fungal elements of lichen, and such famous Fungi as Saccharomyces, Aspergillus, Candida and Neurospora, as well as morels, truffles and similar delicacies. The current understanding is that supposed pre-Devonian even Proterozoic!) lichens are probably artifacts, making the earliest known ascomycote of Carboniferous age. The Ascomycota are the largest and most diverse group of Fungi. They include the yeasts, most of the fungal elements of lichen, and such famous Fungi as Saccharomyces, Aspergillus, Candida and Neurospora, as well as morels, truffles and similar delicacies. The current understanding is that supposed pre-Devonian even Proterozoic!) lichens are probably artifacts, making the earliest known ascomycote of Carboniferous age.
The Ascomycota are united by the presence of asci see glossary entry). Like Basidiomycota, ascomycotes remain indefinitely in the dikaryon state, with the fungal filaments (hyphae) partitioned into cells each containing two haploid nuclei -- one from each parent. Also as in basidiomycotes, nuclear fusion (karyogamy) occurs only in connection with the formation of sexual spores. At that time the newly diploid nucleus undergoes one (sometimes more) round of mitosis, followed by meiosis, to yield eight (or a multiple of eight) haploid nuclei. The nuclei are then partitioned by internal membranes into individual ascospores. The Ascomycota also share with Basidiomycota the use of conidia for the development of asexual spores.
Another unique character (but not present in all ascomycotes) is the presence of Woronin bodies on each side of the septa separating the hyphal segments. The septae are pierced by pores which allow most cytoplasmic constituents (but not nuclei) to travel freely between hyphae. However, if an adjoining hypha is ruptured, the Woronin bodies block the pore to prevent loss of cytoplasm into the ruptured compartment.
For more information, see Ascomycota.
ATW041118. Text public domain. No rights reserved. |
|
ATW??????, page revised MAK120115.

