|
|
Ascomycota |
| Fungi |
Ascomycota - 1 |
The Ascomycota - 1
 The Ascomycota are the largest and most diverse group of Fungi. They include the yeasts, most of the fungal elements of lichen, and such famous Fungi as Saccharomyces, Aspergillus, Candida and Neurospora, as well as morels, truffles and similar delicacies. The current understanding is that supposed pre-Devonian even Proterozoic!) lichens are probably artifacts, making the earliest known ascomycote of Carboniferous age.
The Ascomycota are the largest and most diverse group of Fungi. They include the yeasts, most of the fungal elements of lichen, and such famous Fungi as Saccharomyces, Aspergillus, Candida and Neurospora, as well as morels, truffles and similar delicacies. The current understanding is that supposed pre-Devonian even Proterozoic!) lichens are probably artifacts, making the earliest known ascomycote of Carboniferous age.
Ascomycetes are united by the presence of asci see glossary entry). Like Basidiomycota, ascomycetes remain indefinitely in the dikaryon state, with the fungal filaments (hyphae) partitioned into cells each containing two haploid nuclei -- one from each parent. Also as in basidiomycotes, nuclear fusion (karyogamy) occurs only in connection with the formation of sexual spores. At that time the newly diploid nucleus undergoes one (sometimes more) round of mitosis, followed by meiosis, to yield eight (or a multiple of eight) haploid nuclei. The nuclei are then partitioned by internal membranes into individual ascospores. The Ascomycota also share with Basidiomycota the use of conidia for the development of asexual spores.
ATW041118. Text public domain. No rights reserved.
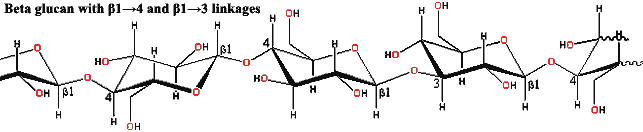 Other than the ascus, not a great deal unites the Ascomycota. Almost all ascomycetes are terrestrial or parasitic. However, a few have adapted to marine or freshwater environments. The cell walls of the hyphae are variably composed of chitin and β-glucans, just as in Basidiomycota. However, these fibers are set in a matrix of glycoproteins containing the sugars galactose and mannose, rather than xylose and mannose as in Basidiomycota [4].
Other than the ascus, not a great deal unites the Ascomycota. Almost all ascomycetes are terrestrial or parasitic. However, a few have adapted to marine or freshwater environments. The cell walls of the hyphae are variably composed of chitin and β-glucans, just as in Basidiomycota. However, these fibers are set in a matrix of glycoproteins containing the sugars galactose and mannose, rather than xylose and mannose as in Basidiomycota [4].
The mycelium of ascomycetes is usually made up of septate hyphae. However, there is 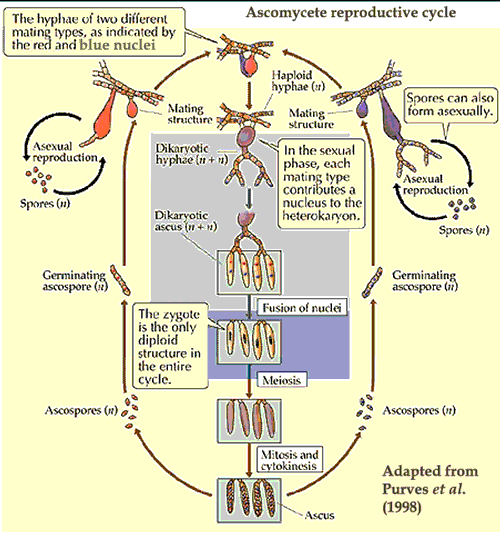 not necessarily any fixed number of nuclei in each of the divisions. The septal walls have septal pores which provide cytoplasmic continuity throughout the individual hyphae. Under appropriate conditions, nuclei may also migrate between septal compartments through the septal pores.
not necessarily any fixed number of nuclei in each of the divisions. The septal walls have septal pores which provide cytoplasmic continuity throughout the individual hyphae. Under appropriate conditions, nuclei may also migrate between septal compartments through the septal pores.
A unique character of the Ascomycota (but not present in all ascomycetes) is the presence of Woronin bodies on each side of the septa separating the hyphal segments which control these septal pores. If an adjoining hypha is ruptured, the Woronin bodies block the pores to prevent loss of cytoplasm into the ruptured compartment. The Woronin bodies are spherical, hexagonal, or rectangular membrane bound structures with a crystalline protein matrix. More derived ascomycetes may have, in addition, septal pore organelles which isolate portions of the mycelium which are undergoing sexual reproduction.
Mycologists long ago learned the same trick which has more recently been discovered by television programming executives. The secret is this: regardless of phylogenetic position, reproductive behavior and physiology lend themselves to the creation of strong and attractive images which enable one to go on for hours without really having to communicate a great deal of substance. In addition, ascomycetes, like the characters in situation comedies, seem to lead sex lives of utterly unreasonable complexity.Sadly, the resemblance ends there, or mycology would be considerably more popular than it is. Quite aside from issues of physical aesthetics, Fungi simply lack the native ability to make us identify with their unrequited yearnings for hyphae of the opposite mating type. Even such dramatic events as ballistospory quite common among ascomycetes) cannot disguise the fact that this is, fundamentally, a rather tedious business once we get past the usual colorful diagram.
At any rate, we have now provided a suitably edifying chart with which to illumine the dark night of ignorance, etc. So, our work here is done; and we may return to matters of more intrinsic interest, secure in the knowledge that our duty to a century of mycological tradition has been fully performed.
It is worth noting that ascomycetes tend to have special relationships with insects. Some are "farmed" by beetles and social insects, while others are parasitic on insects. Yet others produce powerful toxins which are relatively specific to insects. Since both insects and ascomycetes are largely terrestrial and both experienced their first major radiation in the Mississippian, it may be that they shared some significant resource or niche. However, the great 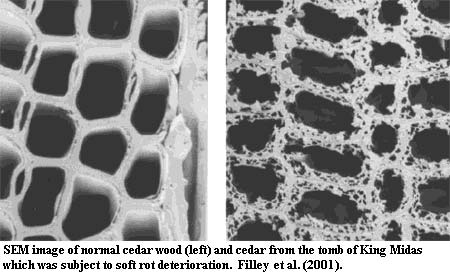 expansion of insect life probably took place in the Serpukhovian, quite late in the Mississippian, and was most closely related to the evolution of flight.
expansion of insect life probably took place in the Serpukhovian, quite late in the Mississippian, and was most closely related to the evolution of flight.
Another notable ecological link is the relationship between ascomycetes and plant life. Both basidiomycetes and ascomycetes developed the ability to digest plant tissues early on. This was a critical development, since the explosive spread of land plants in the Late Devonian and Mississippian was burying atmospheric carbon dioxide at a ferocious rate and increasing oxygen to the point that it became -- explosive. Basidiomycetes and ascomycetes evolved a sort of division of labor in handling this important bit of recycling. Basidiomycetes do most of the heavy lifting by digesting cellulose (brown rot) and lignin (white rot). However, ascomycetes do their bit by digesting the other glucans which hold the other plant materials together (soft rot). The effects of soft rot are shown -- from a most unusual source -- in the image. Filley et al. (2001). Thus the ascomycetes are important in making plant tissues accessible to attack by basidiomycetes. It seems we have the fungi to thank for saving us from a world caught in an unstable alternation between frozen wastes and exploding forests.
Links: Ascomycota; Mycology - Structure and Function - Wall Composition; 561- Lecture#13
ATW041231. Text public domain. No rights reserved.
It is now generally accepted that Ascomycota and Basidiomycota are sister clades, since they share a number of synapomorphies, including septate mycelia, a dikaryotic stage in the life cycle, plectenchymatous structures associated with spore production, and conidia. The only weak spot in this reasoning is that the yeasts, which are rather basal ascomycetes, are secondarily (probably) unicellular and lack some of these multicellular synapomorphies. However, both molecular and morphological comparisons place the yeasts comfortably within the Ascomycota, and most workers are willing to accept that they derived from a yet more basal multicellular ascomycete ancestor with the appropriate characteristics. A certain amount of this sort of thing is likely, given that Ascomycota has a ghost lineage of about 60 My, i.e., the time between the first known appearance of Basidiomycota and the first clear remains of Ascomycota.
The ascomycetes were traditionally divided into the Hemiascomycetes and the Euascomycetes, sometimes with the addition of a probably paraphyletic basal group called Archaeascomycetes. Bold 1973). More recently, mycology was seduced by the spell of the
 Svengalis of Sequencing, with the usual disastrous abandonment of useful nomenclature. We often resist using molecular nomenclature because it is only useful when discussing sequence data and seems to represent a willful rejection of everything else. Like the "Newspeak" of George Orwell's 1984, molecular nomenclature tends to make certain thoughts impossible by denying them a convenient linguistic point of reference. However, at least for the Fungi, we acknowledge the superior pressure of competing considerations, discussed elsewhere. For those reasons we have adopted the new words. For those who wish to mentally translate:
Svengalis of Sequencing, with the usual disastrous abandonment of useful nomenclature. We often resist using molecular nomenclature because it is only useful when discussing sequence data and seems to represent a willful rejection of everything else. Like the "Newspeak" of George Orwell's 1984, molecular nomenclature tends to make certain thoughts impossible by denying them a convenient linguistic point of reference. However, at least for the Fungi, we acknowledge the superior pressure of competing considerations, discussed elsewhere. For those reasons we have adopted the new words. For those who wish to mentally translate:
Taphrinomycotina = Archaeascomycetes
Saccharomycotina = Hemiascomycetes
Pezizomycotina = Euascomycetes
In even older literature, the ascomycetes, as we presently understand them, were divided between two groups, the Ascomycota and the Deuteromycota. The latter were often referred to as fungi imperfecti, i.e., species of fungi for which no sexual stage was known. It is now known that these “imperfect fungi”, formerly Class Hyphomycetes in the Deuteromycota, are anamorphs asexual forms) of the ascomycetes [5].
ATW041231. Text public domain. No rights reserved.
As mentioned above, the Ascomycota are very unevenly divided into Taphrinomycotina, Saccharomycotina, and Pezizomycotina.
 This is almost certainly a paraphyletic basal radiation of ascomycetes. Some may even be more closely related to true yeasts than to pezizomycotines. Bullerwell et al. (2003). Typical members include Neolecta, the fission yeast Schizosaccharomyces, the human pathogen Pneumocystis carinii, and the plant pathogen Taphrina. As might be expected, body forms are particularly diverse among the Taphrinomycotina, ranging from relatively complex filamentous forms to single-celled yeast-like growths.
This is almost certainly a paraphyletic basal radiation of ascomycetes. Some may even be more closely related to true yeasts than to pezizomycotines. Bullerwell et al. (2003). Typical members include Neolecta, the fission yeast Schizosaccharomyces, the human pathogen Pneumocystis carinii, and the plant pathogen Taphrina. As might be expected, body forms are particularly diverse among the Taphrinomycotina, ranging from relatively complex filamentous forms to single-celled yeast-like growths.
Generally, two simple body plans are found for the production of asci: uncovered asci with unicellular growth, or a rudimentary ascocarp with hyphal growth. That is, these fungi grow either unicellularly or with a sparse mycelium, and their sexual phase normally produces naked asci directly from the ascogenous cells. Only Neolecta produces a (primitive) enclosing ascocarp. Liu & Hall (2004).
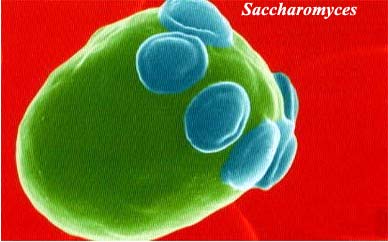 These are the true yeasts, including the Saccharomyces cereviseae of college genetics classes, and the human pathogen Candida. Domestication of Saccharomyces for fermentation of beer goes back at least 9000 years, and probably predates human cultivation of most other plants and animals. This group appears to be monophyletic and to have abandoned multicellularity independently of the yeast like genera in Taphrinomycotina. Basal members of Saccharomycotina still retain a mycelial growth habit. However, none of the Saccharomycotina produces an ascocarp.
These are the true yeasts, including the Saccharomyces cereviseae of college genetics classes, and the human pathogen Candida. Domestication of Saccharomyces for fermentation of beer goes back at least 9000 years, and probably predates human cultivation of most other plants and animals. This group appears to be monophyletic and to have abandoned multicellularity independently of the yeast like genera in Taphrinomycotina. Basal members of Saccharomycotina still retain a mycelial growth habit. However, none of the Saccharomycotina produces an ascocarp.
Saccharomyces became the first eukaryote to have its genome completely sequenced in 1996. Shortly afterwards, a dozen other saccharomycotine genomes were partially sequenced, creating the first opportunity to look seriously at the comparative genomics and evolution of an entire eukaryotic class. So far as we know, this remains the only study of this scope as of this date (041231). The results of this analysis were reported in a series of papers published in FEBS Letters in 2000, all of which are now available at the Génolevures website. See, especially, Gaillardin et al. (2000), Llorente et al. (2000a), Llorente et al. (2000), and Malpertuy et al. (2000). A brief diversity summary, such as the present section, is not the place to take on such a massive body of data. However, we cannot resist the temptation to mention a few of the gems to be found in this gigantic dragon-hoard of information.
First, it appears that yeast evolution is driven by a dynamic balance between gene duplication and deletion. It appears that genes are constantly being shuffled around the genome, with chromosome fragments frequently being duplicated and inserted elsewhere, often as additions to the ends of chromosomes -- the peculiarly labile regions near the telomeres. In most cases, the resulting copies are simply deleted. However, over geological time, the entire genome is shuffled with considerable regularity.
Second, the number of orthologs (copies, but not necessarilly exact copies) of each gene is more stable than one might expect from this mode of evolution. It seems that unnecessary gene copies are deleted rather quickly. This conclusion is also fortified by the observation that genes involved in unused metabolic pathways are almost entirely absent. Thus, for example, Saccharomyces, which feeds solely by fermentation, has no genes orthologous to those coding for enzymes involved in oxidative metabolism.
Finally, and most significantly, yeasts show a strong correlation between the age of a gene and its stability.So, for example, the ancient genes coding for proteins involved in the basic work of transcription and translation are quite stable. By contrast, genes related to cell wall synthesis -- functions unique to the fungi and to particular groups within the fungi -- change quite quickly. This particular study needs to be done at a much higher level of resolution, but the tentative conclusion one may draw is that long-term selective pressures are a significant part of the evolutionary picture. If so, this data may settle an important and contentious point about the process of evolution.
CONTINUED ON NEXT PAGE
