Oxymonadida
| Abbreviated Dendrogram
|
Contents
Summary
Oxymonadida
Polymastigidae |
Taxa on This Page
-
Oxymonadida
-
Polymastigidae
|
This page is about the Oxymonadida, a group of protists. Protists are eukaryotic organisms with only one cell. Protists evolved from the bacteria approximately one billion years ago. Unlike bacteria, protists are eukaryotic cells, the same kind of cell that makes up the bodies of all animals and plants. Eukaryotic cells have several characteristics that are quite different from bacterial cells. The two most important eukaryotic features are the nucleus and the cytoskeleton. The nucleus is a membrane sac inside the cell which contains the cell's DNA the DNA of bacteria is not walled off from the rest of the cell). The cytoskeleton is a system of rigid rods (microtubules) and flexible filaments (microfilaments) made of protein.
Oxymonads are a small group of protists who live inside the digestive tract of termites and other wood-eating insects. Oxymonads probably help break down wood to a more digestible form. However, their exact role is unknown. The oxymonads, in turn, are the hosts for several species of bacteria who also play a part in digesting wood particles. Oxymonad cells are often completely covered in a layer of long, slightly spiral bacteria call "spirochetes." Several lines of evidence suggest that the oxymonads developed a "partnership" with these bacteria long before oxymonads began to live inside insects.
Oxymonads are distinguished by the presence of a large axostyle, a long bundle of microtubules that extends almost the entire length of the cell. In some cases, it actually protrudes from the end of the organism. Oxymonads typically also have a pelta, a sheet of microtubules which covers the end of the cell containing the nucleus. The nucleus is located at one end of the cell which, by convention we call the "anterior." There are typically four flagella also located at the anterior end.
Many workers believe that oxymonads may be the closest living relatives of the very first protists. Oxymonads lack mitochondria and have no Golgi apparatus. Almost all eukaryotic cells have these structures. Unfortunately, these are both primitive traits and "absence" characters. It is generally a mistake to classify organisms on the basis of some supposed primitive condition, or on the basis of features they don't have. So, for example, human beings and birds share, with most fishes, the absence of a hard shell and a "pulley" system for the jaw muscles. That doesn't mean that humans are either closely related to birds or that humans are more fish-like and "primitive" than turtles, who have both features. This problem became obvious recently when both molecular and structural evidence showed that oxymonads are closely related to Trimastix, a protist which does have structures similar to mitochondria.
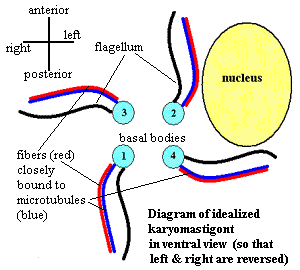 However, there may be better reasons to think that oxymonads are close to the root of the Eukarya. Oxymonads, Trimastix, and several other groups of protists all share a very distinctive group of linked structures called the karyomastigont. A diagram of an idealized karyomastigont is shown on the right. No known organism has a karyomastigont of this form. It is shown this way simply so that we can understand its basic symmetry and as a speculation about how it might have been arranged in the very earliest protists. The karyomastigont is built around four basal bodies. Each basal body gives rise to a flagellum. In addition, each basal body is associated with a "microtubule root," a place where a sheet or bundle of microtubules originates. Each group of microtubules is, in turn, bound to a fiber (not microfilaments, but a different kind of microfiber). Each of the basal bodies, and their associated structures, is oriented in a different direction. At least one of the basal bodies is also connected to the nucleus in a fashion which, it seems, is not understood. However, there may be better reasons to think that oxymonads are close to the root of the Eukarya. Oxymonads, Trimastix, and several other groups of protists all share a very distinctive group of linked structures called the karyomastigont. A diagram of an idealized karyomastigont is shown on the right. No known organism has a karyomastigont of this form. It is shown this way simply so that we can understand its basic symmetry and as a speculation about how it might have been arranged in the very earliest protists. The karyomastigont is built around four basal bodies. Each basal body gives rise to a flagellum. In addition, each basal body is associated with a "microtubule root," a place where a sheet or bundle of microtubules originates. Each group of microtubules is, in turn, bound to a fiber (not microfilaments, but a different kind of microfiber). Each of the basal bodies, and their associated structures, is oriented in a different direction. At least one of the basal bodies is also connected to the nucleus in a fashion which, it seems, is not understood.
Prof. Lynn Margulis, in particular, has argued that the karyomastigont represents the evolutionary remains of an ancient mobility symbiosis which gave rise to the first eukaryotic cells. Whether or not this is correct, the basic structural features of many apparently primitive protists can be understood in terms of variations on this basic theme. ATW030825. |
 Oxymonadida:
Oxymonadida:
Range: The Oxymonadida have no fossil record. The oxymonads are known largely as a component of the complex community of bacteria and protists which live in the hindgut of wood-eating insects, mainly termites. The oxymonads are one of three protist groups typically found in this environment, the others being trichomonads and hypermastigids [M+98]. No oxymonad has been successfully cultured in isolation [M+98], which creates some severe technical obstacles to detailed study.
The exact nature of the relationship with the insect host is not understood [K+98]. It seems likely that the oxymonads are involved in the breakdown of lignin [K+98]. It may be that their main metabolic interaction is with bacterial commensals, rather than directly with the insect host. Oxymonads are frequently found with numerous spirochete bacteria attached to what may be specific binding sites. Although some bacterial species are associated only with particular protist species in the hindgut, there does not appear to be any simple 1 : 1 relationship [N+03]. Some bacteria also have preferential binding areas on the surface of the protist, but most apparently do not [N+03]. In addition, as one might expect, the make-up of these mixed protist - bacterial communities varies considerably between termite species [N+03]. The specific relationships between bacterium and protist are said to be far more stable than the bacterium
- termite relationships [N+03].
Phylogeny: *: Polymastigidae + Oxymonadidae + (Saccinobaculidae + Pyrsonymphidae + Streblomastigidae))). The phylogenetic position of the Oxymonadida has been quite labile, although the general tendency has been to place this taxon well toward the root of the Eukarya because of the absence of both mitochondria and Golgi apparatus [S+02]. Until quite recently, many workers accepted Cavalier-Smith's assignment of the group to a Metamonada, including oxymonads, Diplomonada, and Retortamonada [S+02] [M+03]. These were assigned to the paraphyletic subkingdom Archezoa, within an equally paraphyletic Kingdom Protozoa [C98]. However, recent work, both molecular and morphological, has indicated a close relationship between oxymonads and Trimastix, an excavate genus [D+01] [S+02] [SS+02]. The morphological basis for this assignment is discussed below, in connection with the cytoskeleton of the Polymastigidae. The putative relationship with Trimastix is troublesome because Trimastix is not amitochondriate. It lacks true mitochondria, but appears to have organelles resembling hydrogenosomes [D+01].
As of this date (8/03) no one has yet worked out a sensible-looking tree which would accommodate the new data. However, it would not be surprising if this the new tree looked rather similar to the conventional wisdom of 50 years ago, which also had these groups relatively close together. We note (with irritating smugness) that the Trimastix - oxymonad connection is reasonably compatible with our current best guess phylogeny. The relationship is compatible -- not because we have abandoned Metamonada, like the Molecule Masters -- but because we have abandoned Excavata. In our Best Guess phylogeny, the ventral feeding groove and its homologues are primitive for Eukarya.
A somewhat similar view is taken by Margulis et al. [M+00]. These workers postulate that the Eukarya evolved by genetic fusion of a sulfur-metabolizing archaean with its motility symbiote, a spirochete-like eubacterium. We will not review their theory in detail here, but merely point to one of its consequences: the fundamental importance of the karyomastigont. This is the most general and primitive form of a complex of the nucleus with (usually four) basal bodies and associated flagella, microtubular tissues and fibers. We will begin to examine this complex below in connection with the polymastigids and the critical work of Simpson et al. [S+02]. Part of this complex involves a recurrent flagellum lying in a groove lined by two fibers, one rather amorphous and one ordered and striated, each with a fixed relation to one of the basal bodies. That description fits both the feeding groove of the Excavata and a much less conspicuous structure in oxymonads. Structures like the axostyle and pelta, notable synapomorphies of oxymonads turn out, on closer inspection, to be probable hypertrophies of other elements of this basic karyomastigont apparatus. Whether or not [M+00] have correctly identified the source of the karyomastigont, it does appear to be a very distinctive structure common to all groups at the base of the Eukarya.
Another, perhaps less meaningful phylogenetic clue comes from molecular studies using the sequence of elongation factor 1α [M+98]. Phylogenies based on EF-1α suggest a relationship (probably not a sister clade relation) between oxymonads and diplomonads, with both groups relatively near each other and near the base of the Eukarya [DR98] [M+98] [M+01].
At the moment (8/03) we take no detailed position on the external relationships of the oxymonads, although we strongly favor the connection with the excavates, as discussed. Within Oxymonadida, we follow -- more or less -- the scheme of Moriya et al. [M+03].
Characters: General: Mostly flagellates, all known species are commensals usually in intestines of termites & other insects able to live on lignin.
Peripheral structures: The cell surface is naked. However, the cells be covered with ectosymbiotic bacteria [M+03].
Membranes: The cell membrane may bear receptors of an unknown kind for attachment of commensal bacteria. There are no extrusomes and the membranes are otherwise naked.
 Motility organs: four flagella in two pairs.
Motility organs: four flagella in two pairs.
Cytoskeleton: four basal bodies arranged in two separated pairs and giving rise to several major microtubular roots, some with associated nonmicrotubular roots. The two pairs of basal bodies are "maintained apart" by a preaxostyle and associated fiber [B91]. The basal bodies also give rise to a paracrystalline organelle, the axostyle [B91], made up of multiple sheets of microtubules created or recruited by the preaxostyle [B91]. The axostyle may be able to undulate. That is, in some species, the microtubular sheets are able to slide past one another. The axostyle and preaxostyle[B91], are said to be synapomorphies of the Oxymonadida. However a similar structure has long been known in the diplomonad Trichomonas vaginalis, a human venereal disease agent, and related parasitic forms [L+03]. The α-tubulins of these two groups do not seem to be particularly closely related. Moriya et al. [M+01] attempt to distinguish the Trichomonas axoneme as being non-motile. However, Tritrichomonas foetus, which causes venereal disease in cattle, has a style of motility (described as a "rolling, jerky motion") which at least causes one to speculate that the axostyle is involved in its motility.
Mitochondria: Mitochondria and related structures are absent [S+02] [C99].
Other organelles: Golgi apparatus or dictyosomes are also absent [S+02] [M+03]. Oxymonad rRNA has larger hypervariable regions than the rRNA of other Eukarya, including their putative close relative, Trimastix [M+03]. In particular, [M+03] state that an expansion of stem 43 in the V4 region may be synapomorphic for Oxymonadida.
Nuclei: typically with one nucleus, but may be multinucleate [M+03].
Reproduction: Like most protists, oxymonads are facultatively sexual [DR98]. For oxymonads, as for middle-aged humans, "sex is both infrequent and occurs in response to an environmental stimulus." [DR98: 779].
Links: Microscope: Oxymonadida; Untitled Document.
References: Brugerolle 1991) [B91]; Cavalier-Smith (1998) [C98]; Cavalier-Smith 1999) [C99]; Dacks & Roger 1998) [DR98]; Dacks et al. 2001) [D+01]; Kudo et al. (1998) [K+98]; Lecke et al. 2003) [L+03]; Margulis et al. 2000) [M+00]; Moriya et al. 1998) [M+98]; Moriya et al. 2001) [M+01]; Moriya et al. 2003) [M+03]; Silberman et al. 2002) [SS+02]; Simpson et al. (2002) [S+02]. ATW030816.
Polymastigidae: Chilomitus, Cochlosoma, Monocercomonoides, Paranotila, Polymastix.
Range: no fossil record. Found in gut of insects and vertebrates as commensals or normal fauna. None are known to be parasitic.
Phylogeny: Oxymonadida: (Oxymonadidae + Saccinobaculidae + (Pyrsonymphidae + Streblomastigidae))) + *. Phylogeny based largely on [M+03]. The basal position of the polymastigids within Oxymonadida seems to be uncontroversial. See, e.g., [B91].
Characters: Small flagellates. The characters below referred to [S+02] pertain to Monocercomonoides, and may not apply to the other genera of this group. It may be worth noting that Monocercomonoides is not a typical oxymonad. If nothing else, at 5µ in length it is the smallest known member of the taxon [M+03].
Peripheral structures: Polymastigids lack both a holdfast and a rostellum. Consequently, they are free-swimming gut commensals [M+03].
Motility organs: Polymastigids bear 4 flagella, at least one (or only one [B91]) of which is recurrent, in two pairs [K66]. All flagella bear an acroneme. The recurrent flagellum adheres to the body for some distance and then continues as a free whip, to which is attached a funis is this distinct from the acroneme?). One group (BB1 & BB2) is placed ventrally, the other more dorsally, in the anterior part of the cell (BB3 & BB4) [S+02]. Note that, historically, the BB1-2 group was referred to as "dorsal." We have reversed the dorsoventral terminology for the reasons given by [S+02], as discussed below (or perhaps above, as the case may be). The two pairs of basal bodies are connected by a preaxostyle as discussed below.
Cytoskeleton: An axostyle or axial filament is present, but slender [K66]. A row (or sheet?) of microtubules or pelta covers the anterior end. The pelta is closely associated with a microtubular root (R2) which originates near the BB3-4 complex. It is attached to the ventral basal bodies. Two separate groups of basal bodies are connected by a 'U'-shaped fiber, the preaxostyle. The preaxostyle appears as a broad, curved sheet of microtubules which face the nucleus [B91]. It is most strongly associated with BB1 [S+02]. On the side facing away from the nucleus, the preaxostyle adheres to a 50 nm fiber with a latticework appearance [S+02] [B91]. The preaxostyle serves as an site of microtubule nucleation for the axostyle [B91].  A short, hook-like fiber, the 'H' fiber' emerges from the edge of the preaxostyle, close to BB1. The H fiber extends posteriorly briefly, following the edge of a shallow channel in which the proximal portion of flagellum 1 rests [S+02]. The H fiber is broad, sheet-like and bears striations with a periodicity of 30 nm [S+02]. The opposite edge of the channel is associated with a sheet of microtubules (R1) also originating near BB1[S+02]. Near BB1, R1 is associated with an amorphous fiber which rapidly diminishes posteriorly [S+02]. A lone pair of microtubules originate between the preaxostyle and BB1 and run down the center of the ventral channel [S+02].
A short, hook-like fiber, the 'H' fiber' emerges from the edge of the preaxostyle, close to BB1. The H fiber extends posteriorly briefly, following the edge of a shallow channel in which the proximal portion of flagellum 1 rests [S+02]. The H fiber is broad, sheet-like and bears striations with a periodicity of 30 nm [S+02]. The opposite edge of the channel is associated with a sheet of microtubules (R1) also originating near BB1[S+02]. Near BB1, R1 is associated with an amorphous fiber which rapidly diminishes posteriorly [S+02]. A lone pair of microtubules originate between the preaxostyle and BB1 and run down the center of the ventral channel [S+02].
Simpson et al. [S+02] assert that Monocercomonoides and, by extension all oxymonads, are morphologically related to excavate taxa, particularly Trimastix. To understand the comparison, we are compelled to detour into the structure of Trimastix, which will serve as a morphotype for the excavate body plan.  Trimastix is shown here in ventral view, laboriously adapted from [OK+99]. As one might expect of an excavate, the ventral face is dominated by a large ventral feeding groove. The kinetosome complex of Trimastix is in an anteroventral position. For convenience, the size of the kinetosome elements has been greatly exaggerated in the diagram. The complex contains four basal bodies, inserting into four flagella: BB1 posterior, BB2 anterior, BB3 right, and BB4 left [S+02]. Sadly, we must keep careful track of the numbers for purposes of judging homology.
Trimastix is shown here in ventral view, laboriously adapted from [OK+99]. As one might expect of an excavate, the ventral face is dominated by a large ventral feeding groove. The kinetosome complex of Trimastix is in an anteroventral position. For convenience, the size of the kinetosome elements has been greatly exaggerated in the diagram. The complex contains four basal bodies, inserting into four flagella: BB1 posterior, BB2 anterior, BB3 right, and BB4 left [S+02]. Sadly, we must keep careful track of the numbers for purposes of judging homology.
The complex also produces four microtubule roots and three fibers [S+02]. BB2 is associated with the origin of the anterior microtubular root which, in turn, produces a "dorsal fan" or "dorsal spray" of secondary microtubules in the anterior part of the cell [S+02]. BB1 is associated with three microtubular roots [S+02]. The large left and right ventral roots originate on either side of BB1[S+02]. These microtubules define the margins of the ventral groove and close the groove where they converge again posteriorly [OK+99]. A singlet root also emerges posteriorly from between BB1 and the right ventral root [S+02]. The 'I' fiber covers the ventral face of the right root. It has a distinctive latticework structure [S+02]. The 'B' fiber is sheet-like in most taxa, bearing 30 nm striations. It supports the right wall of the feeding groove anteriorly [S+02]. The 'C' fiber performs the same function on the left side. The singlet root follows and defines the floor of the groove [S+02].
| Excavate-Oxymonad Homologies According to Simpson et al. (2002) |
| Trimastix |
Monocercomonoides |
| Ventral feeding groove |
Ventral flagellar channel |
| Basal Body 1 |
Basal Body 1 |
| Basal Body 2 |
Basal Body 4 |
| Anterior root |
R2 |
| dorsal fan |
pelta |
| Right ventral root |
preaxostyle |
| Left ventral root |
R1 |
| I fiber |
preaxostyle fiber |
| B fiber |
H fiber |
| C fiber |
amorphous fiber |
| Singlet root |
doublet root |
Given these features, the argument for homology seems compelling. The primary differences are in the direction of the I fiber (preaxostyle). In excavates, the I fiber helps support the right side of the feeding groove. In oxymonads, it wanders off in the opposite direction and joins the two pairs of basal bodies.
Simpson et al. [S+02] assume without much discussion that Trimastix and the excavates are the plesiomorphic form. We question whether the matter of polarity quite so easily resolved. The ventral feeding groove is a highly specialized structure. We are also dealing with a question of very deep time. Billions of generations separate today's genera from the last common ancestor of Trimastix and the oxymonads. The morphology of their common ancestors will have to be deduced. We are unlikely to find it wandering about loose.
Mitochondria: as in all oxymonads, there are no mitochondria or related structures.
Nuclei: The nucleus is anterior and lies behind the preaxostyle and the wall of the pelta.
Links: Polymastigidae.
References: Brugerolle 1991) [B91]; Kudo (1966) [K66]; Moriya et al. 2003) [M+03]; O'Kelly et al. 1999) [OK+99]; Simpson et al. (2002) [S+02]. ATW030816.
ATW061129
last revised ATW070103, edited RFVS111206
checked ATW061130
