Alveolata
 Alveolata is a high-order group of Eukarya whose principal members include (a) the Ciliophora (e.g., Paramecium), (b) a large group of revolting parasites called the Apicomplexa (e.g. Plasmodium, the organism responsible for malaria), and (c) the dinoflagellates, a hugely successful group of marine photosynthetic organisms. In addition, the Haplosporidia may fall within the Alveolata. For the moment, however, we will leave them out of the mix.
Alveolata is a high-order group of Eukarya whose principal members include (a) the Ciliophora (e.g., Paramecium), (b) a large group of revolting parasites called the Apicomplexa (e.g. Plasmodium, the organism responsible for malaria), and (c) the dinoflagellates, a hugely successful group of marine photosynthetic organisms. In addition, the Haplosporidia may fall within the Alveolata. For the moment, however, we will leave them out of the mix.
Despite their considerable success, the Alveolata are apparently a taxon of relicts. They are united by the presence of small vesicles (alveoli) in, or just under, the plasma membrane. The function of the alveoli is unknown, although they are believed to form part of a complete inner membrane system. The usual speculation is that they function in ion transport and in structural stabilization of the cell membrane. The outer membrane is pierced by micropores of unknown function. Siddall et al. (2001). The internal membrane systems may be present, but dictyosomes are often reduced. The implication, we assume, is that the alveoli are taking up the transport function normally assumed by the Golgi apparatus. The Microscope site adds: "Flagella when present (whether as flagella or cilia) typically with at least one cross-striated fibrous root". We take this character to be primitive for Eukarya. The Alveolata share with their sister clade, Chromista, an interesting flagellar accessory known as the mastigoneme,which is described at the glossary entry. The Alveolata also go in for other strange, and sometimes unique, organelles, but these are not synapomorphies of the whole taxon, and we will take them up as need be. All of the Alveolata prefer oxygen-rich environments and engage in oxidative metabolism using mitochondria of the usual, tubulocristate kind.
But that's about the extent of their similarities. The three alveolate taxa otherwise seem very different. The ciliates are free-living heterotrophs, most of which inhabit soils. The dinoflagellates are marine (benthic or planktonic) photosynthetic autotrophs. The Apicomplexa are all obligate parasites. Nevertheless, these three disparate strands consistently braid together in both molecular and morphological tests. We are therefore forced to the conclusion that we're looking at a relict taxon, in which only isolated fragments remain of an originally continuous spectrum of diversity.
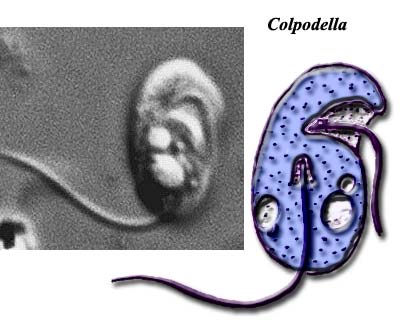
Image credit: Colpodella images both from Microscope.
ATW041031. Text public domain. No rights reserved.
Alveolate Phylogeny: Colpodella, Parvilucifera, and Perkinsus
 In this context, the recent work of Siddall et al. (2001) is particularly welcome. The Siddall group addressed the molecular phylogeny of three, very similar, alveolate genera with debatable affinities: Colpodella, Perkinsus, and Parvilucifera. The thought was that these forms might represent missing links between the three major alveolate taxa.
In this context, the recent work of Siddall et al. (2001) is particularly welcome. The Siddall group addressed the molecular phylogeny of three, very similar, alveolate genera with debatable affinities: Colpodella, Perkinsus, and Parvilucifera. The thought was that these forms might represent missing links between the three major alveolate taxa.
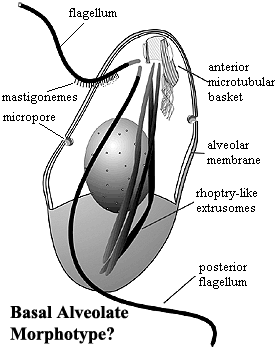
The basic structure of these forms is shown in the figure from Siddall et al. (2001). The essential structures are not really very different from the karyomastigont structure which has been proposed as the basal organelle arrangement of the Eukarya. We see a complex anterior microtubular array from which the flagella emerge. There are fibrous sheets radiating from the "mastigont" as well, although these are not as closely integrated with the flagellae as in Metamonada. Instead of only a posterior flagellar/feeding groove, we see both posterior and lateral grooves. At least one flagellum bears mastigonemes. The nucleus is no longer bound into the mastigont. The internal, alveolar membrane system is present, with micropores associated with the alveoli. When displayed in this manner, the system is curiously reminiscent of the polar filament coils of the Microsporidia
-- a similarity which may not be coincidental if a secondary membrane system should turn out to be basal to Metabiotiformes. Extrusomes (another apparent synapomorphy of Metabiotiformes) are present and take the form of elongate, somewhat club-shaped sacs terminating anteriorly. These are quite similar to the rhoptries of the Apicomplexa. Similarly, all genera have some sort of anterior conoid-like structure associated with predation or intracellular insertion in the Apicomplexa.
Siddall's theoretical approach to molecular work is outstanding. It is constrained by morphological data and gives appropriate attention to the morphological implications. Therefore his results ought to have rather higher credibility than many such studies -- particularly here, where morphology will probably not be sufficient to resolve phylogeny. The Siddall group also wisely uses two completely different gene sequences: for actin and for SSU rRNA. They do not find a unique solution, but their results tend to yield a phylogeny along the lines shown in the image.
 The Siddall group's execution of this particular study has, however, been heavily criticized for various technical errors. See, e.g., Cavalier-Smith & Chao (2004). These criticisms are almost certainly correct with respect to Colpodella. Nonetheless, the differences between the ultimate results of Siddall and Cavalier-Smith are not particularly earth-shattering at the level of resolution relevant here. Neither Siddall nor Cavalier-Smith were ultimately able to say with any confidence just where Colpodella lies. The answer to that question may well depend on which species and isolate one studies. Perkinsus and Parvilucifera are Dinozoa in both studies. The apparent disagreement comes only from the usual (we were about to say "pig-headed", but exerted our usual iron self-discipline just in time) refusal of protistologists to adopt reasonable phylogenetic definitions.
The Siddall group's execution of this particular study has, however, been heavily criticized for various technical errors. See, e.g., Cavalier-Smith & Chao (2004). These criticisms are almost certainly correct with respect to Colpodella. Nonetheless, the differences between the ultimate results of Siddall and Cavalier-Smith are not particularly earth-shattering at the level of resolution relevant here. Neither Siddall nor Cavalier-Smith were ultimately able to say with any confidence just where Colpodella lies. The answer to that question may well depend on which species and isolate one studies. Perkinsus and Parvilucifera are Dinozoa in both studies. The apparent disagreement comes only from the usual (we were about to say "pig-headed", but exerted our usual iron self-discipline just in time) refusal of protistologists to adopt reasonable phylogenetic definitions.
Curiously, none of the three genera turns out to be a definite apicomplexan. Parvilucifera and Perkinsius are Dinozoa. Colpodella turns out to be "a paraphyletic mess hovering about the base of Miozoa." C. Taylor, pers. comm. 2004. Since none of these genera show any obvious synapomorphies with the three main alveolate clades, Siddall et al. propose that these three collectively exemplify the basal type of all Alveolata. From our previous discussion, this observation may be limited just to the Miozoa. Still, it looks to be a pretty good bet for that group. Other than micropores and alveoli, the basic structure looks not so different from, for example, an oxymonad.
In the fullness of time, we hope to get more deeply into this particularly weird corner of phylospace. Certainly there is much more here than just green dinoflagellates and evil, parasitic sporozoans. As in human society, there are any number of odd characters who are impossible to categorize -- as well as an unsettling number of evil, parasitic green dinoflagellates and beneficial sporozoans.
ATW041031. Text public domain. No rights reserved. Revised ATW041104. This section has benefited considerably from the thoughtful comments of Christopher Taylor, Univ. of Auckland who, of course, is not responsible for any thoughtless errors we have made in applying them.
Alveolata |
| |
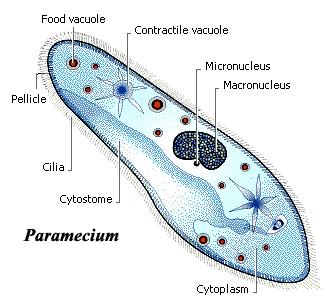 The Ciliophora are the ciliates, including the ubiquitous Paramecium of high school biology texts. They may have a fossil history going back into the Precambrian, if chitinozoans are, as suspected, ciliophoran coverings. Ciliophora are both very common and quite diverse. Most are free-living aquatic predators. They may be benthic or planktonic and, at times, Ciliophora may account for a very large fraction of standing plankton biomass. The Ciliophora are the ciliates, including the ubiquitous Paramecium of high school biology texts. They may have a fossil history going back into the Precambrian, if chitinozoans are, as suspected, ciliophoran coverings. Ciliophora are both very common and quite diverse. Most are free-living aquatic predators. They may be benthic or planktonic and, at times, Ciliophora may account for a very large fraction of standing plankton biomass.
The Ciliophora are characterized, reasonably enough, by the presence of cilia. Generally speaking, cilia are simply short flagella. At some stage of life, ciliophorans have numerous cilia covering some substantial fraction of the cell membrane. The cilia often occur in rows (kineties), which helps to explain how hundreds of separate cilia can be coordinated in locomotion. The ciliary bases are attached to the pellicle, a peripheral cytoskeleton. In addition, Ciliophora have an epiplasm composed of a fibrous mesh.
The plasma membrane has both a permanent cytostome "mouth" and a permanent
cytoproct "anus." The cytoproct is a feeding groove presumably derived from the old posterior flagellar groove. The more exterior portion of the cytostome is underlain by a fibrous network, as was the ancestral flagellar groove. the groove terminates in a region which packages the food particles into digestive vacuoles and pinches them off into the cytoplasm. The kineties around the cytostome are arranged to funnel particles deeper into the feeding groove. Also associated with the plasma membrane are extrusomes, which rapidly eject short threadlike structures (as do the possibly homologous micronemes of Apicomplexa). These extrusomes function in predation, defense, and in forming cysts in various Ciliophora.
 The cytoplasm contains, in addition to digestive vacuoles, contractile vacuoles which probably function in the control of osmotic pressures. They can open to the medium and presumably discharge excess internal water or ions. The cytoplasm contains, in addition to digestive vacuoles, contractile vacuoles which probably function in the control of osmotic pressures. They can open to the medium and presumably discharge excess internal water or ions.
There are two types of nuclei, micronuclei and macronuclei, either of which may be present singly or in several copies. The micronuclei are diploid, with condensed chromatin. They appear to function largely in reproduction. Sexual reproduction is common and, in some species, required for long-term survival. During sexual reproduction a cytoplasmic bridge is constructed between the two cells, and micronuclei are exchanged over this bridge. In this process, the macronuclei simply break down. The macronuclei appear to contain multiple copies of particular genes needed for day-to-day metabolic functions.
Links: Protozoa B; Introduction to the Ciliata.
ATW041031 Text public domain. No rights reserved. Revised ATW041104. |
|
Miozoa was originally erected by Cavalier-Smith to unite Apicomplexa and Dinozoa. It seems that he is no longer happy with that name and has attempted to substitute "Myzozoa" = "sucking life." And so it does on occasion, but that's no reason to arbitrarily unseat the senior name. Consequently we retain the older name -- at least until our usual cowardice in such matters results in the more usual fawning capitulation to taxonomic fashion. Cavalier-Smith & Chao (2004: 194) characterize the taxon as "[p]redominantly haploid, typically uninucleate alveolates with zygotic meiosis; lacking separate macronuclei; ancestrally and typically with two centrioles and cilia only; anterior cilium often with simple hairs. Trichocysts typically with a dense basal rod that is square in cross section and a less dense distal region composed of hollow twisted tubules. When trichocysts are present cortical alveoli are typically inflated and morphologically discrete, often with internal plates; when trichocysts are absent they are typically highly compressed and often fused into an inner membrane complex. Myzocystosis [sic --> myzocytosis] and/or rhoptries and micronemes are very widespread, and possibly even ancestral." This is somewhat unhelpful since it is largely a description of the alternative character states of Dinozoa and Apicomplexa. In fact, of all of the characters mentioned which we can clearly identify, all are either plesiomorphic "ancestral" -- not unique to Miozoa) or are apomorphies of included taxa (only apply to some Miozoa). No synapomorphies are identified.
ATW041031 Text public domain. No rights reserved. Revised ATW041104. |
| |
Dinozoa was originally created by Cavalier-Smith to contain the Dinoflagellates and the "Protalveolata." The latter are an artificial group of misfits, such as the three genera studied by Siddall et al. 2001). Since Miozoa is indisputably a crown group, it only makes sense to treat Dinozoa and Apicomplexa as the corresponding stem groups. Thus Dinozoa = dinoflagellates > sporozoans. There is no room for a "Protalveolata," even if such a group existed. As matters stand, Perkinsus and Parvilucifera are dinozoans. Most other well-known dinozoans are traditional dinoflagellates, and these are discussed elsewhere. A variety of other strange and wonderful creatures also inhabit this phylospace which we will have to get to another day.
Image Credit: Xiphoridium from Andrew McRae's Dinoflagellates pages.
ATW041031 Text public domain. No rights reserved. Revised ATW041104. |
|
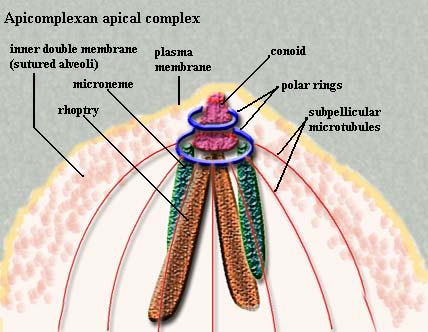 The Apicomplexa are common parasites of insects, vertebrates, and almost everything else. They are characterized by a particularly fiendish parasitic tool-kit called the apical complex (hence, of course, the name). Added to this, apicomplexans have a thick, triple outer layer which is very flexible, but nearly impervious to biological, and even to most chemical, agents. In some cases it is possible to clean out the entire cytoplasm with detergents and still recover -- more or less intact -- the reinforced membrane structure. The Apicomplexa are common parasites of insects, vertebrates, and almost everything else. They are characterized by a particularly fiendish parasitic tool-kit called the apical complex (hence, of course, the name). Added to this, apicomplexans have a thick, triple outer layer which is very flexible, but nearly impervious to biological, and even to most chemical, agents. In some cases it is possible to clean out the entire cytoplasm with detergents and still recover -- more or less intact -- the reinforced membrane structure.
The outer layer of the apicomplexan cell is an ordinary(?) plasma membrane. However, this membrane is buttressed by an inner, double membrane made up of flattened alveoli sutured together. Finally, the whole business is reinforced with the cellular equivalent of rebar -- longitudinal bundles of microtubules running from the apical complex back towards the posterior end of the cell. These are cross-linked in some fashion.
The membrane is broken only by a simple cytostome consisting of an invagination of the plasma membrane. It is very similar in structure to the flagellar grooves that are occur in many other protist groups.
The components of the apical complex rhoptries, micronemes,
polar rings, the conoid, and subpellicular microtubules) are described in the appropriate glossary entries. We defer further discussion to a time when we can go beyond brief summaries. Apicomplexans have life cycles which are complex. In fact they would seem almost comical were it not for the fact that they are so efficient, and so often deadly to the host species. The basic life cycle may be said to start when an infective stage, or sporozoite, enters a host cell, and then divides repeatedly to form numerous merozonts. Some of the merozonts transform into reproductive cells, or gamonts. Gamonts join together in pairs and form a gamontocyst. Within the gamontocyst, the gamonts divide to form numerous gametes. Pairs of gametes then fuse to form zygotes, which give rise by meiosis to new sporozoites.
Motile forms of Apicomplexa crawl along the substratum in a non-amoeboid fashion known as gliding motility, which is poorly understood. Many apicomplexan species have flagellated gametes.
Links: Apicomplexa; Introduction to the Apicomplexa; Sporozoa Notes.
ATW041102. Text and image public domain. No rights reserved. Revised ATW041104. |
checked ATW061220, edited RFVS111206

