| Palaeos: Paleozoic | Ordovician Period | |
| Ordovician period | Ordovician |
| Page Back | Back: Cambrian | Back: Neoproterozoic | Up: Paleozoic | Unit Home |
| Page Next | Next: Silurian | Next: Mesozoic | Down: Early Ordovician | Time |
 The Ordovician Period is the second period of the Paleozoic Era. This important period saw the origin and rapid evolution of many new types of invertebrate animals which replaced their Cambrian predecessors. Primitive plants move onto land, until then totally barren. The supercontinent of Gondwana drifted over the south pole, initiating a great Ice Age that gripped the earth at this time. The end of the period is marked by an extinction event.
The Ordovician Period is the second period of the Paleozoic Era. This important period saw the origin and rapid evolution of many new types of invertebrate animals which replaced their Cambrian predecessors. Primitive plants move onto land, until then totally barren. The supercontinent of Gondwana drifted over the south pole, initiating a great Ice Age that gripped the earth at this time. The end of the period is marked by an extinction event.
The Ordovician System of strata was founded by Lapworth in 1879 to resolve the Murchison-Sedgwick conflict over their overlapping claims for their Silurian and Cambrian systems. Benton & Harper (1997). The name "Ordovician" comes from an ancient Celtic tribe that once inhabited the region in Wales where rock strata of this period occur.
The Ordovician was originally divided into two epochs, Bala and Dyfed. More recently, the Tremadoc was removed from the Cambrian and a three-fold division of Ordovician strata instituted.
Image: An Ordovician drama - an endocerid cephalopod captures a trilobite. From Earth History Resources.
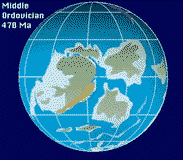 During the Ordovician, Southern Europe, Africa, South America, Antarctica and
Australia remained joined together into the supercontinent of Gondwanaland,
which had moved down to the South Pole. North
America straddled the equator, and was about 45 degrees clockwise from
its present orientation. Western and Central Europe were separate
from the rest of Eurasia, and were rotated about 90 degrees counterclockwise
from their present orientation, and was in the southern tropics.
North America is engaged in a slow collision with the microcontinent of
Baltica, which forms the core of what is later to become Europe.
The Iapetus Ocean continues to shrink as the previously passive margins
of Baltica and North America converge. Where the Iapetus was, mountains
are thrust up, remnant strata of which remain today in Greenland, Norway,
Scotland, Ireland and north-eastern North America. Scotland and England
are united into a single landmass.
During the Ordovician, Southern Europe, Africa, South America, Antarctica and
Australia remained joined together into the supercontinent of Gondwanaland,
which had moved down to the South Pole. North
America straddled the equator, and was about 45 degrees clockwise from
its present orientation. Western and Central Europe were separate
from the rest of Eurasia, and were rotated about 90 degrees counterclockwise
from their present orientation, and was in the southern tropics.
North America is engaged in a slow collision with the microcontinent of
Baltica, which forms the core of what is later to become Europe.
The Iapetus Ocean continues to shrink as the previously passive margins
of Baltica and North America converge. Where the Iapetus was, mountains
are thrust up, remnant strata of which remain today in Greenland, Norway,
Scotland, Ireland and north-eastern North America. Scotland and England
are united into a single landmass.
 The
Ordovician is, from the point of view of stratigraphers, the Age of Graptolites.
Graptolites are thought to be the remains of hemichordates of some kind, and
often look like a cross between a conodont and a crumpled pipe-cleaner. Like
conodonts, their remains are common and correlate reasonably well over broad
areas of the world. The base of the Ordovician is the level at which
planktonic (thus widespread) graptolites are first found. The end of the
Ordovician (i.e., the base of the Silurian)
corresponds to an extinction which eliminated all but one graptolite genus
[3].
The
Ordovician is, from the point of view of stratigraphers, the Age of Graptolites.
Graptolites are thought to be the remains of hemichordates of some kind, and
often look like a cross between a conodont and a crumpled pipe-cleaner. Like
conodonts, their remains are common and correlate reasonably well over broad
areas of the world. The base of the Ordovician is the level at which
planktonic (thus widespread) graptolites are first found. The end of the
Ordovician (i.e., the base of the Silurian)
corresponds to an extinction which eliminated all but one graptolite genus
[3].
In the last century, the gold standard in Ordovician stratigraphy was the British system. If nothing else, the Ordovician System of Britain, with its wonderful Welsh names, dated back to the dawn of geology. However, after intense study by the International Commission on Stratigraphy, it became depressingly clear that the whole thing would have to be redone if there was to be any hope of worldwide correlation for the Ordovician.
As a result, all of the age names have now been changed (except the Tremadoc) and given more rigorous definitions. However, outside the world of stratigraphers, the scientific literature still primarily uses the old names. Hence, we provide some general equivalences. Note that a few of the approximate British "equivalents" are very approximate indeed. In particular, the Arenig included not only the Floian and the Dapingian, but also the first ~2 My of the Darriwilian. Mitchell et al. (1997).
Note that the current (080216) versions of the Wikipedia entries on the Ordovician show a different, internationalized, version of the British system, in which the base of the Llanvirn is dragged down to coincide with the base of the Middle Ordovician. In this version, the Llanvirn occupies almost all of the Middle Ordovician except for a brief Llandeilo Age at the end. There is no room in this system for the Arenig, as it was usually understood, and the Wikipedia entries consequently avoid the use of that term.
Our representation of the British system is also bastardized, but in a different way. The term Llandeilo is indeed still frequently found, and the Llandeilo was one of the original Ordovician series, lying between the Llanvirn and Caradoc (i.e. at the top of the Middle Ordovician). In many versions of British Ordovician stratigraphy, the Llandeilo was merged with Llanvirn even before the ICS started playing with the whole system. This is the representation we have adopted, because it allows us to show the Arenig as it was used during most of the XXth century. Accordingly we avoid using the term Llandeilo. This discussion may give you a good understanding of why the ICS thought it better to start all over again when it began to address Ordovician stratigraphy.
| Period | Epoch | Age | Approx. British |
Base |
Duration |
Silurian |
Llandovery | Rhuddanian | 443.7 | 4.7 | |
Ordovician |
Late Ordovician |
Hirnantian | Ashgill |
445.6 | 1.9 |
| Katian | 455.8 | 10.2 | |||
Caradoc |
|||||
| Sandbian | 460.9 | 5.1 | |||
Middle Ordovician |
Darriwilian | Llanvirn (& Llandeilo) | 468.1 | 7.2 | |
Arenig |
|||||
| Dapingian | 471.8 | 3.7 | |||
Early Ordovician |
Floian | 478.6 | 6.8 | ||
| Tremadoc | Tremadoc |
488.3 | 9.7 | ||
Cambrian |
Furongian | Cambrian X | 492.0 | 3.7 |
 A major transgression in the Middle Ordovician created widespread shallow, warm epicontinental
seas. Thus, most of the Ordovician was favorable for marine life,
particularly around the well-studied European and North American cratons.
However, the Ordovician ended in a brief (300-500 ky), but severe, ice
age. Gondwana,
particularly Africa, straddled the
South Pole and
became extensively glaciated. There were even glaciers in what is now the Sahara.
Metazoans were severely effected. About 60% of animal genera became extinct,
making this the second or third most deadly mass extinction of the Phanerozoic [1].
A major transgression in the Middle Ordovician created widespread shallow, warm epicontinental
seas. Thus, most of the Ordovician was favorable for marine life,
particularly around the well-studied European and North American cratons.
However, the Ordovician ended in a brief (300-500 ky), but severe, ice
age. Gondwana,
particularly Africa, straddled the
South Pole and
became extensively glaciated. There were even glaciers in what is now the Sahara.
Metazoans were severely effected. About 60% of animal genera became extinct,
making this the second or third most deadly mass extinction of the Phanerozoic [1].
 As
a natural consequence, a good deal of attention has been focused on the causes
of the Ordovician Ice Age. In fact, it is not easy to see how an ice age
could have occurred. Atmospheric carbon dioxide levels are believed to
have been 8 to 20 times their current values. This ought to have prevented
anything approaching an ice age. Sea levels were high through most of the
Ordovician. They dropped, dramatically (about 50 m), in connection with
the ice age, but it is hard to tell whether this was cause, effect, or
both. One independent factor which would affect both pCO2 and
sea level is the rate of sea floor spreading along mid-ocean ridges. As we
might expect, the length of well-established mid-ocean ridges, i.e., the ridge
between Gondwana, to the south, and Baltica plus Laurentia, to the north, was
unusually short during the Late Ordovician. A former ridge between the two
northern continents became inactive about this time. However, there may
have been a very long ridge to the Northwest of Laurentia. The information
is too sparse to be certain. In any event, the absence of active ocean
crust formation would only affect the rate of CO2 outgassing, not the
rate at which it was locked away in sediments.
As
a natural consequence, a good deal of attention has been focused on the causes
of the Ordovician Ice Age. In fact, it is not easy to see how an ice age
could have occurred. Atmospheric carbon dioxide levels are believed to
have been 8 to 20 times their current values. This ought to have prevented
anything approaching an ice age. Sea levels were high through most of the
Ordovician. They dropped, dramatically (about 50 m), in connection with
the ice age, but it is hard to tell whether this was cause, effect, or
both. One independent factor which would affect both pCO2 and
sea level is the rate of sea floor spreading along mid-ocean ridges. As we
might expect, the length of well-established mid-ocean ridges, i.e., the ridge
between Gondwana, to the south, and Baltica plus Laurentia, to the north, was
unusually short during the Late Ordovician. A former ridge between the two
northern continents became inactive about this time. However, there may
have been a very long ridge to the Northwest of Laurentia. The information
is too sparse to be certain. In any event, the absence of active ocean
crust formation would only affect the rate of CO2 outgassing, not the
rate at which it was locked away in sediments.
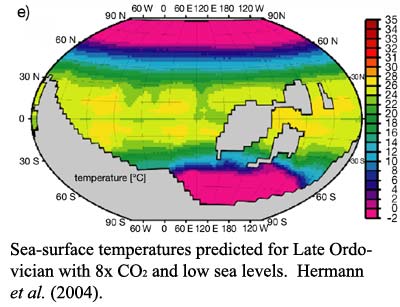
The state of the art in mathematical modeling of the problem is described in the recent work of Hermann et al. (2004). The results are frustratingly uninformative. Sea surface temperatures for the later Ordovician are extremely sensitive to atmospheric carbon dioxide levels and to not much else. Even the changes in geography, which brought more land surface close to the South Pole, seem to have little effect on the outcome. The result in the image indeed predicts glaciers in Gondwana at the end of the Late Ordovician. Unfortunately, it also predicts glaciers in Gondwana at the beginning of the Late Ordovician, even with much higher sea levels. Thus, if the models have anything to tell us, it is that there must have been a very strong draw-down of CO2 over the Late Ordovician. No one, at this point, has been able to offer evidence suggesting a plausible agent which would remove roughly half of all atmospheric carbon dioxide in 10-15 My.
ATW041124. Text public domain. No rights reserved.
The Ordovician was an age of evolutionary experimentation, in which new organisms evolve to replace those that died out at the end of the Cambrian. It was also one of the largest adaptive radiations in the Earth's history. The number of families of known marine invertebrates (mostly hard shelled forms, as - soft-bodied types rarely left fossils) increases from about 200 at the end of the Cambrian to around 500 in the early Ordovician. The widespread shallow, warm continental seas were the perfect environment for many groups of organisms. Micro-organisms such as colonial blue-green algae - stromatolites - are widespread. Foraminifera (marine amoebas which build tiny shells) evolve for the first time. Organisms called acritarchs, although existing during the Precambrian, become more common. Stromatoporoids (possibly sponge-like organisms) also appear.
 An interesting phenomenon is the sudden increase
in filter feeding organisms. Cambrian animals were predominately crawling
mud-grubbers and detritivores with a few swimming and burrowing predators
thrown in. Filter feeders (such as Dinomischus and Lepidocystis)
although an important part of the fauna, had not been exceedingly common.
In the Ordovician an increase in the amount of micro-plankton would be
the obvious explanation for the sudden increase in number and diversity
of filter-feeders. Groups absent or under-represented in the Cambrian suddenly become more important. We see the first appearance of the corals,
including both rugose and tabulate forms, bivalve molluscs, and the planktonic graptolites (creatures
distantly related to vertebrates). [note: the orange anemone-like
animals in top-right of the underwater
scene at the top of this page are rugose corals, possibly Streptelasma
or Lambeophyllum.].
An interesting phenomenon is the sudden increase
in filter feeding organisms. Cambrian animals were predominately crawling
mud-grubbers and detritivores with a few swimming and burrowing predators
thrown in. Filter feeders (such as Dinomischus and Lepidocystis)
although an important part of the fauna, had not been exceedingly common.
In the Ordovician an increase in the amount of micro-plankton would be
the obvious explanation for the sudden increase in number and diversity
of filter-feeders. Groups absent or under-represented in the Cambrian suddenly become more important. We see the first appearance of the corals,
including both rugose and tabulate forms, bivalve molluscs, and the planktonic graptolites (creatures
distantly related to vertebrates). [note: the orange anemone-like
animals in top-right of the underwater
scene at the top of this page are rugose corals, possibly Streptelasma
or Lambeophyllum.].
 The lophophorates (animals that suck food from the water using a special fringe of tentacles
around the mouth) are more filter feeders that do well in the Ordovician.
The Bryozoa appear in
large numbers, and constitute the most predominant colonial animals of
the time. Each bryozoan polyp is a tiny creature, not unlike a coral
animal. Their distant cousins the hard-shelled brachiopods
are also successful. After humble Cambrian beginnings the articulate
brachiopods greatly increase in diversity and abundance, with no fewer
than fourteen new superfamilies. The Orthid
and Strophomenid
orders were especially diverse.
The lophophorates (animals that suck food from the water using a special fringe of tentacles
around the mouth) are more filter feeders that do well in the Ordovician.
The Bryozoa appear in
large numbers, and constitute the most predominant colonial animals of
the time. Each bryozoan polyp is a tiny creature, not unlike a coral
animal. Their distant cousins the hard-shelled brachiopods
are also successful. After humble Cambrian beginnings the articulate
brachiopods greatly increase in diversity and abundance, with no fewer
than fourteen new superfamilies. The Orthid
and Strophomenid
orders were especially diverse.
Among the molluscs were newcomers such as bivalves, which were not common during this time, although the Furongian Archaeogastropods develop at a moderate pace. A much more spectacular success story were the nautiloid cephalopods. Small and rare in the Furongian, the nautiloids evolve quickly along many different lines. At least ten different orders flourished at this time, all but one appearing for the first time during the early or middle part of the Ordovician. This astonishing diversity included straight-, curved, loosely coiled, and tightly coiled shelled types, and even one group (the Ascocerids) that in order to become lighter and more streamlined lost the a large part of their shell altogether. These intelligent carnivorous molluscs replaced the Cambrian Anomalocarids as the dominant life form and top predator of the world's ocean. The biggest, such as the endocerids (see graphic at top of page), attained huge size; with shells of upto 10 meters in length they were the largest animal that, up until that time, had ever lived.
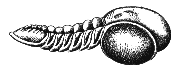 Ordovician
trilobites were for the most part quite different from their Cambrian predecessors. Many evolved bizarre spines and nodules, others, such as Aeglina prisca (left) from the warm shallow seas of what is now Bohemia, were clearly pelagic nektonic swimming forms, and developed huge eyes with over a thousand facets, while still others went the other direction and lost their eyes altogether. Some trilobites developed shovel-like snouts for ploughing through mud, others fused the segments of their bodies, while the curious Trinucleids developed a broad pitted margin around the head shield.
Ordovician
trilobites were for the most part quite different from their Cambrian predecessors. Many evolved bizarre spines and nodules, others, such as Aeglina prisca (left) from the warm shallow seas of what is now Bohemia, were clearly pelagic nektonic swimming forms, and developed huge eyes with over a thousand facets, while still others went the other direction and lost their eyes altogether. Some trilobites developed shovel-like snouts for ploughing through mud, others fused the segments of their bodies, while the curious Trinucleids developed a broad pitted margin around the head shield.
 A
number of early echinoderm
experiments die out, others straggle on, while still others increase in
diversity. The echinoderms included both a great many stalked (filter-feeding)
and a few mobile (some predatory) forms. Among the bizarre forms
were the carpoids, which were able to push themselves along the mud by
means of a stout "tail". It has even been suggested that certain
carpoids are ancestral to the vertebrates,
A
number of early echinoderm
experiments die out, others straggle on, while still others increase in
diversity. The echinoderms included both a great many stalked (filter-feeding)
and a few mobile (some predatory) forms. Among the bizarre forms
were the carpoids, which were able to push themselves along the mud by
means of a stout "tail". It has even been suggested that certain
carpoids are ancestral to the vertebrates,  but in view of the
Furongian
vertebrate origin this is unlikely. During the Ordovician the Crinoids,
rare during the Cambrian, suddenly appear and diversify in large numbers.
Like the brachiopods these sessile benthic (attached bottom-living) invertebrates
were to become an important group of filter feeders throughout the rest
of the Paleozoic.
but in view of the
Furongian
vertebrate origin this is unlikely. During the Ordovician the Crinoids,
rare during the Cambrian, suddenly appear and diversify in large numbers.
Like the brachiopods these sessile benthic (attached bottom-living) invertebrates
were to become an important group of filter feeders throughout the rest
of the Paleozoic.
The Ordovician was the high point of the graptolites. These colonial hemichordates possessed an anatomical structure suggestive of a portion of a spinal cord. They were vertebrate cousins rather than vertebrate ancestors. They evolved from benthic attached Furongian forms and diversified in a number of different planktonic types, including single-branched single and double rowed colonies, two-branched, four-branched, and even spiral forms.
The vertebrate ostracoderms remain rare, although several different groups of Pteraspidomorphi evolve. Their cousins the conodont animals, worm-like or eel-like organisms known mostly from numerous isolated denticles (which were used to support some kind of grasping or breathing structure in the mouth or throat) represent a major component, quite possibly predators and certainly nektonic/pelagic, in the marine food-chain.
Finally, a humble start to a new adventure: during the Ordovician the first creeping lichens and hepatophytes move onto land, the beginning of a great new experiment of life.
Image: The graptilite Tetragraptus from Prehistoric Life (the Victoria Museum).
The isolated and drifting continents serve as island arks, on the continental shelves of which marine organisms engage in evolutionary experiments. There are two main geographical provinces, a northern, equatorial tropical one, and a southern cool-water one centered around Gondwanaland. In the case of trilobites, North America and northwestern margins of Europe, Spitzbergen, Siberia and north-east Russia are characterized by a diverse fauna that H. B. Whittington calls the Bathyurid fauna. This includes trilobites of the families Bathyuridae, Hystricuridae, Asaphidae, Komaspididae, Remopleurididae and Pliomeridae. A subprovince around Baltica bathyurids and pliomerids are rare and asaphids belong to different genera, indicated the Iapetus Ocean remained wide enough to act as a barrier to migration for these shallow water continental-shelf forms. In the cooler waters of Gondwanaland are found the Selenopeltis and Hungaiid-Calymenid faunal provinces, including trilobites of the family Hungaiidae, Calymeniidea, Pliomeridae, Illaenidae, and endemic Asaphidae. The differences between Bathyurid fauna and Hungaiid-Calymenid faunal provinces was due to a combination of climate factors and geographical separation.
The same biogeographical distribution seems to apply to articulate brachiopods as well. The Balto-Scandian region is better defined in the brachiopod then the trilobite faunas, and for much of the period can be considered a distinct biogeographical province.
In the later part of the Ordovician genera of families previously limited to one faunal province appear in another, indicating a tendency towards migration and cosmopolitism. Changed in oceanic circulation (distribution of planktonic larvae etc) along with approaching continental masses would have made possible migrations of shallow water benthos.
The tremendous evolutionary radiation of nautiloid cephalopods meant an increase in the level of consciousness in Paleozoic oceans. Cephalopods are the most intelligent of all invertebrates, owing to the development of elaborate manipulative organs (tentacles for touching and grasping). One researcher described the common octopus as like a sort of aquatic dog or cat. Although Ordovician cephalopods were probably not equal to modern forms in intelligence, they were certainly superior to that of contemporary (non-cephalopod) life-forms.
| Page Back | Page Up | Page Next |