| Palaeos: Paleozoic | Middle Ordovician Epoch | |
| Ordovician period | Middle Ordovician |
| Page Back | Back: Early Ordovician | Back: Cambrian | Up: Ordovician | Unit Home |
| Page Next | Next: Late Ordovician | Next: Silurian | Down: Dapingian | Time |
The Middle Ordovician includes the Dapingian (Middle Arenig) and Darriwilian (Late Arenig, Llanvirn and Llandeilo) Ages. Historically, it has been the most difficult and contentious boundary in the Ordovician. The Ordovician conveniently left us good records of both graptolites and conodonts, as well as trilobites, brachiopods, and some stable isotope excursions. Unfortunately, the non-Gondwanan continents were scattered and fragmented during much of the period. As a result, the fauna of one continent often had little or no overlap with the fauna of another, making correlation extremely difficult. This is particularly hard luck because the precise stratigraphy of the Middle Ordovician lies at the core of a key issue. As we will see, a completely unreasonable number of important clades first appeared in this epoch, during what has been called the "Great Ordovician Biodiversity Event." This event was the mirror image of a mass extinction, and paleontologists are naturally anxious to understand precisely when, and under what conditions, such an unusual radiation might have occurred.
Special Credit: Thanks to Prof. Stan Finney for responding to a conodont issue. We rewrote the stratigraphic problem considerably after receiving his explanation. ATW080307.
 The base of the Middle Ordovician is the most difficult
stratigraphic problem we
have looked at thus far.
Eventually, after some false starts, the ICS selected, as the Middle Ordovician global stratotype section and point, the
Huanghuachang Section, an exposure of the lower Dawan Formation near
Yichang, in Hubei Province. Wang et al.
(2005). This lies in
Central China, near the Yangtze River, and -- by no coincidence at all -- just a
few km up the road from the GSSP for the Hirnantian
Age of the Late Ordovician.
Oddly, the
Huanghuachang Section lacks a good set of graptolite index fossils. This is
unfortunate for an Ordovician GSSP, since so much Ordovician stratigraphy has
been done using graptolites. However, there are at least a few graptolite
markers, and a number of other index fossils are present.
The base of the Middle Ordovician is the most difficult
stratigraphic problem we
have looked at thus far.
Eventually, after some false starts, the ICS selected, as the Middle Ordovician global stratotype section and point, the
Huanghuachang Section, an exposure of the lower Dawan Formation near
Yichang, in Hubei Province. Wang et al.
(2005). This lies in
Central China, near the Yangtze River, and -- by no coincidence at all -- just a
few km up the road from the GSSP for the Hirnantian
Age of the Late Ordovician.
Oddly, the
Huanghuachang Section lacks a good set of graptolite index fossils. This is
unfortunate for an Ordovician GSSP, since so much Ordovician stratigraphy has
been done using graptolites. However, there are at least a few graptolite
markers, and a number of other index fossils are present.
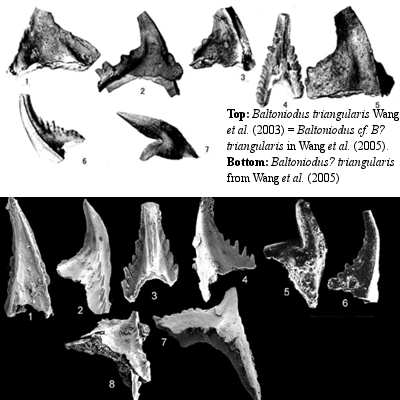
The original guide event for the Middle Ordovician was supposed to be the first appearance of the conodont Tripodus laevis on the theory that T. laevis would permit correlation with Ordovician conodont zones in Northern Europe, Newfoundland, the Western U.S., and elsewhere. Unfortunately, for reasons too complicated to get into here, T. laevis has been described under at least four different names, including Acodus combsi, Scolopodus alatus, and Tripodus combsi. In the profoundly understated words of Bergstr�m & Albanesi (2001), "the fact that different authors use a different species designation for the same biozonal index species is likely to cause confusion not only among non-conodont workers." To make matters worse, someone pointed out -- during an ICS field trip to a proposed GSSP, no less -- that T. laevis seemed to appear at different levels even within related exposures in Nevada. Thus, when Wang et al. (2003) proposed using another conodont, Baltoniodus triangularis [5], and the well-sampled Chinese section at Huanghuachang, the idea was well-received. Even better, the lowest B. triangularis level coincided almost exactly with the appearance of T. laevis, which was also present in this section.
But the Middle Ordovician seems to be cursed. We suspect that this is the evil work of a secret cabal of immortal druids, justly enraged at the abandonment of the old Welsh names for the Ordovician stages. Whatever the ultimate cause, further investigation showed that the fossils described as Baltoniodus triangularis at Huanghuachang, were in fact two different, overlapping species, neither one of which was actually Baltoniodus triangularis. Indeed, neither species had ever been properly described or named. In the iconography of stratigraphy, these are referred to as Baltoniodus? triangularis and Baltoniodus? cf. B? triangularis. We may refer to the original conodont and its two doppelgangers as Bt, B?t, and B??t.
This problem was not confined to Huanghuachang. As we understand the facts, many of the European Bt zones turned out to be B?t or overlapping bands of Bt and B?t. Unfortunately, the conodont which defined the proposed Huanghuachang GSSP was neither Bt nor B?t, but B??t -- a species apparently unique to the Dawan Formation and thus utterly useless for correlation.
One can only imagine the state of mind of the Ordovician Subcommission, as the members watched 20 years of hard work start to go up in smoke. Fortunately, somone with a very practical turn of mind realized that by deftly moving the proposed GSSP up-section a few meters, it could be made to coincide with the first appearance of B?t, which would then correlate with the numerous wrongly named, but correctly mapped, European sections.
Thus, in Wang et al. (2005), the base of the Middle Ordovician was elevated about six meters from its position in Wang et al. (2003) and a question mark was inserted to redefine the level as the earliest appearance of conodont Baltoniodus? triangularis -- not to be confused with Baltoniodus triangularis (sensu Wang et al., 2003), also known as Baltoniodus? cf. B? triangularis (sensu Wang et al., 2005). A few problems were created by the last-minute change from Bt to B?t. For example, the authors state that, "This [B?t] level is similar to that proposed by Webby (1994, 1998) as a boundary marker and coincides with the base of the Tripodus laevis." Wang et al. (2005: 106). Unfortunately, this is no longer true, as their figures correctly show. See Wang et al. (2005: 107, fig 3).
We are tempted to refer to this epoch as the "Middle? c.f. Ordovician." But this would be more than a little unfair, given the myriad of stratigraphic blowouts and pot-holes succesfully traversed on the way to the Huanghuachang GSSP. Even if we now have a Middle Ordovician GSSP which is slightly ambiguous, we are very much better off than the we were as recently as the 1990's. If there turn out to be real, as well as theoretical, problems with the GSSP, they will come to light soon enough, and the matter can be corrected. ATW080215; rev'd ATW080307.
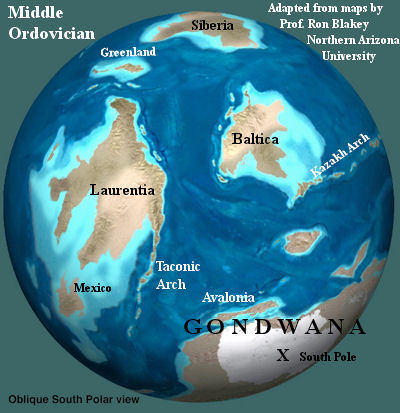 Geography
GeographyAs mentioned, the Middle Ordovician saw the continents scattered. The East coast of Laurentia (North America) would eventually be augmented by two successive waves of refugees from Gondwana, Avalonia and Armorica. However, both were still well-separated from Laurentia during the Middle Ordovician, as were various future parts of southern Baltica (Europe), which also derived from Gondwana. Siberia, Greenland, and Kazakh likewise had little connection to any other landmass. Cocks & Torsvik (2002). The positions and connections of the Chinese and Southeast Asian terranes are even less well understood. They may well have been widely scattered to the east of Baltica, as in the reconstruction by Cocks & Torsvik. However, Australia was probably secured to the main mass of Gondwana (Africa, India, South America, and both Antarcticas) around the South Pole, which was located in today's West Africa.
We should note two disagreements we have with Prof. Blakey's map. First, he shows a south polar ice cap. We do not know what the evidence is for this detail. Several groups have found no evidence of glacial conditions in North Africa at this time. See the discussion of Ordovician climate for more information on the possibility of ice sheets in the Ordovician. Second, the current information on Siberia places it south of the equator, probably due west of Baltica, during the Middle Ordovician.
Because the continents were well-separated, the animals who lived in the shallow waters around them tended to be endemic. That is, the conodonts, mollusks, trilobites, etc. of one continent had little or no genetic contact with those of any other. This makes life difficult for stratigraphers, but also may have furnished more natural laboratories for the "experiments" of evolutionary chance. It also increased the amount of shallow water available for animal life -- and shallow waters were the only environment in which animals lived at this time. ATW080215; rev'd 080311.

One of the defining characteristics of almost all geological epochs is the nature of its reefs. However, during the ~50 My preceding the Middle Ordovician, there had been almost no reefs of any kind. The Middle Ordovician marks the return of the entire reef biome. Stanley (1998).
Like a great deal of Middle Ordovician life, these reefs were a mixture of forms, some old and some very new. These reefs were not extensive, and many were no more than new varieties of bacterial stromatolites reinforced with sponges and calcifying red algae (see image). To these were added crinoids, which had probably first evolved in the Middle Cambrian, as well as tabulate corals and bryozoans, who were Early Ordovician newcomers. However, the Middle Ordovician also saw the development of the first massive stromatoporoid sponges and rugose corals, which together set the pattern for the next 100 or 200 My (depending on exactly how one defines "the pattern"). Benton & Harper (1997); Webby (2004).
As mentioned above, the nearshore, and specifically the nearshore sea bottom, was probably the only environment which supported animal life at the beginning of the Middle Ordovician. That was no longer true at the end of the epoch. First, animals began to colonize deepwater marine environments, as shown by the presence of graptolites and of trace fossils. Hagadorn (1998) (citing work of SC Finney and others).
 Second, open water, planktonic life began to be an important part of animal
ecospace. Peterson
(2005) argues that animals developed large, floating, yolk-filled eggs
several times by the end of the Cambrian. This allowed embryos to disperse
more widely, as they could float for longer periods, feeding on yolk before
settling to the bottom. By the Middle Ordovician, this pelagic picnic was
being harvested by the planktivores, such as the feeding larvae of the
polychaete worms Nereis and Chaetopterus. Id.
Second, open water, planktonic life began to be an important part of animal
ecospace. Peterson
(2005) argues that animals developed large, floating, yolk-filled eggs
several times by the end of the Cambrian. This allowed embryos to disperse
more widely, as they could float for longer periods, feeding on yolk before
settling to the bottom. By the Middle Ordovician, this pelagic picnic was
being harvested by the planktivores, such as the feeding larvae of the
polychaete worms Nereis and Chaetopterus. Id.
Toward the end of the epoch comes the event for which the Middle Ordovician is really well known, the great surge in metazoan diversity of the Darriwilian. This Ordovician "explosion" is best known from the articulate brachiopods, because of their excellent fossil record, and particularly the rhynchonellids. Stanley (1998); Cocks & Torsvik (2003). The most spectacular radiation of the age was among cephalopod mollusks. Several new orders radiated or appeared for the first time in the Middle Ordovician, including the Endocerida, Ellesmerocerida, and the nautiloid Tarphycerina.
 Equally
important are very early vertebrate remains. Several different vertebrate types
seem to have lived at this time. So, for example, isolated fragments of
some
arandaspid-like fishes from the
Amadeus Basin in Australia
and from Bolivia.
Janvier (1996).
In the last few years, new Ordovician thelodonts have also been recovered in
arctic Canada, some of which may be of Darriwilian age.
Sansom & Elliott
(2002). Possible
chondrichthyan
scales have been found in the Harding Sandstone of the western United States (Sansom
et al., 1996) together with
Astraspis and the unclassifiable Eriptychius (Halstead,
1973).
Equally
important are very early vertebrate remains. Several different vertebrate types
seem to have lived at this time. So, for example, isolated fragments of
some
arandaspid-like fishes from the
Amadeus Basin in Australia
and from Bolivia.
Janvier (1996).
In the last few years, new Ordovician thelodonts have also been recovered in
arctic Canada, some of which may be of Darriwilian age.
Sansom & Elliott
(2002). Possible
chondrichthyan
scales have been found in the Harding Sandstone of the western United States (Sansom
et al., 1996) together with
Astraspis and the unclassifiable Eriptychius (Halstead,
1973).
The significance of these particular remains is that they show the development of all but one of the principal types of vertebrate mineralized tissues at a rather early date. Janvier (1996). On theoretical grounds, some writers have proposed that this correlates with an (equally theoretical) large-scale gene duplication event. (There is no question that many chordate regulatory genes were duplicated, but it is less clear whether this was an "event" or a long series of individual duplications). ATW080215.
Image credit: from D&D Fossils & Meteorites.
 This
is the first epoch for which it is meaningful to add this heading.
There is little doubt that plant life had, at last, crept onto the margins of
the land by this time. In fact, the earliest confirmed plant fossils may
be a bit older, from the later Floian. They
consist of the spores of a liverwort-type plant some 476 My old.
Shaw & Renzaglia
(2004). The understanding that they are true land plants is derived
from the co-occurrence of a decay-resistant wall and tetrahedral form which,
together, are diagnostic of land plants.
Kenrick & Crane
(1997).
This
is the first epoch for which it is meaningful to add this heading.
There is little doubt that plant life had, at last, crept onto the margins of
the land by this time. In fact, the earliest confirmed plant fossils may
be a bit older, from the later Floian. They
consist of the spores of a liverwort-type plant some 476 My old.
Shaw & Renzaglia
(2004). The understanding that they are true land plants is derived
from the co-occurrence of a decay-resistant wall and tetrahedral form which,
together, are diagnostic of land plants.
Kenrick & Crane
(1997).
Fossilized fungal hyphae have also been recovered from the latest Darriwilian of Wisconsin, USA. The indications are that these were mycorrhizal fungi: symbiotic fungi which serve as, or are associated with, plant roots. Redecker et al. (2000). This is an important step. It suggests that, by the end of the epoch, plants were growing in regions where they were not covered with water, and therefore required a method of extracting minerals from the soil, at least for substantial periods of time. Thus, by the end of the Middle Ordovician, a hypothetical observer in space might well have seen the beginnings of a pale green fuzz along river valleys and flood plains, far from open water. ATW080215.
| Page Back | Page Top | Page Up | Unit Home | Page Next |
ATW080215.
Last revised ATW080311