Sauropoda: Diplodocomorpha (or Diplodocoidea)
Taxa on This Page
- Apatosaurinae X
- Barosaurus X
- Cetiosauriscus X
- Dicraeosauridae X
- Diplodocidae X
- Diplodocinae X
- Diplodocoidea X
- Diplodocomorpha X
- Diplodocus X
- Nigersaurus X
- Rebbachisauridae X
- Rebbachisaurus X
Rebbachisauridae
 The semi-scientific literature (like these Notes) frequently takes pleasure in nit-picking popular depictions of paleontological material. Perhaps a more interesting and useful exercise might be to worry less about why it isn't accurate. We might instead assume that it is accurate: an accurate representation of some paleocritter unknown to Twenty-First Century science. We might then attempt to determine what it is -- or at least what it isn't. Most science, after all, is not about proving good ideas. We can only disprove bad ones.
The semi-scientific literature (like these Notes) frequently takes pleasure in nit-picking popular depictions of paleontological material. Perhaps a more interesting and useful exercise might be to worry less about why it isn't accurate. We might instead assume that it is accurate: an accurate representation of some paleocritter unknown to Twenty-First Century science. We might then attempt to determine what it is -- or at least what it isn't. Most science, after all, is not about proving good ideas. We can only disprove bad ones.
Consider, for example, the possible (if improbable) sauropod in the figure from Watterson 1989). We might start with the hypothesis that it is some sort of rebbachisaurid. After all, why not? The Rebbachisauridae were founded on a scapula, a few vertebrae in moderately bad shape, and part of a femur. René Lavocat 1954) devoted just over 1 page to these remains in the course of a four-page article on miscellaneous gleanings buried in the deep Sahara. The paper, in turn, was buried in Volume 15 of a 1950-54 series of papers with the stunningly uninformative title: Questions Diverses de Géologie Générale. Not surprisingly, Rebbachisauridae has been used as garbage taxon for various bits and pieces of unidentifiable sauropod from the East End of Nowhere for the last 50 years.
So is it a rebbachisaurid? Clearly not. Its dentition doesn't seem to be arranged as a battery. The nares are on the rostrum and face forward, instead of being above the eyes, facing upward. There are no tall spines on the vertebrae, and the front legs are a bit too short.
Perhaps a diplodocoid? That would make more sense for North America, but it cannot be. Diplodocoid teeth are even more extreme. The diplodocoid occipital condyle faces ventrally, suggesting that the head was held higher and at an angle to the line of the neck. The rostrum is somewhat elongated, but seems too tall for a diplodocoid.
Is it a diplodocomorph at all? Perhaps a nemegtosaur? The quadratojugal might be right for a nemegtosaur, but the basal diplodocomorph synapomorphies are absent. Again, the foliate, grooved teeth do not meet diplodocomorph specs. The anterior jaw line is unclear, but doesn't seem to be square enough. The nares are wrong, as well.
Reluctantly, we conclude that this interesting specimen falls outside the bounds of the Diplodocomorpha. We will have to look elsewhere for an answer. And, while we're at it, what are those hairy little things making all the noise? ATW030127.
Descriptions
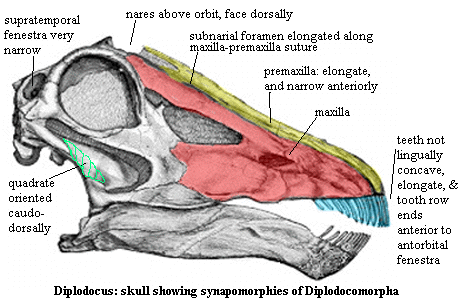 Diplodocomorpha: = Diplodocoidae of Upchurch (1998) =Diplodocoidea, diplodocimorpha, etc.). Diplodocus > Saltasaurus.
Diplodocomorpha: = Diplodocoidae of Upchurch (1998) =Diplodocoidea, diplodocimorpha, etc.). Diplodocus > Saltasaurus.
Long, peg-like teeth, limited to anterior jaw; $ teeth lack lingual concavity; $ anterior jaw "subrectangular"; dentary with angular "chin", anteriorly directed basipterygoid processes; $ transverse narrowing of the rostral end of the premaxilla & premaxilla elongated posteriorly; $ elongation of the subnarial foramen along maxilla - premaxilla suture; extreme retraction of the external nares; $ nares face dorsally, posterior to orbits; $ no internarial bar; anterior extension of the quadratojugal placed beyond the anterior border of the orbit; $ supratemporal fenestra width <10% width of occiput; infratemporal fenestra oval or slit shaped; cervical ribs shorter than associated centra (?); very high and undivided neural spine on dorsal vertebrae with a deep pleurocoel on the centrum; tall neural arch in caudals (at least 1.5x height of centra); wing-like transverse processes in anterior caudals; whip-like tails (30 or more elongate boconvex posterior caudals); broad, paddle-like scapular blade; humerus/femur ratio less than 0.70.
Note: The preferred spelling should be "Diplodocomorpha." The taxon Diplodocimorpha Calvo & Salgado 1995 is a different group, defined by them as Rebbachisaurus tessonei + Diplodocidae. The citation (we have not read the article) is Calvo, JO & L Salgado (1995), Rebbachisaurus tessonei sp. nov. a new Sauropoda from the Albian-Cenomanian of Argentina; new evidence on the origin of the Diplodocidae, Gaia 11: 13-33.
Image: original figure ascribed to Marsh, OC (1896), The Dinosaurs of North America, with coloring and labels added. I'm unsure whether the element in the original is the quadrate or an internal palatal element, so I have sketched in the outline of the quadrate as shown in figures from other sources.
References: Upchurch (1998). ATW030617.
Cetiosauriscus:
Range: Middle Jurassic of England
Phylogeny: Diplodocomorpha : (Rebbachisauridae + Diplodocoidea) + *.
Introduction: Cetiosauriscus stewarti, of Callovian age, is found in the Lower Oxford Clay of Cambridgeshire, England. Cetiosauriscus is the earliest known of the high-hipped and long-necked diplodocomorphs, representing the second of the two primary sauropod feeding strategies. Whereas the giraffe-like camarasaurs and brachiosaurs used their long forelimbs, high shoulders and and necks to reach into vegetation, the diplodocids relied on a different stratergy. They were built like a suspension bridge, with their great arched backbone and long tails putting their centre of gravity over their long hind legs, thus enabling them to rear on their hind legs, using their tail as a support, and so feed on leaves 15 metres above the ground.
In addition to C. stewarti, a "Cetiosauriscus" greppini (Huene 1922) (syn. Cetiosaurus, Ornithopsis) is known from at least 4 partial skeletons without skulls found in the Late Tithonian Unter- Virgula- Schichten of Kanton Bern, Switzerland. These specimens have a humerus measuring about 60 cm, and a femur 70 cm. The overall length about 8.5 metres, the weight about 1 tonne. This was a fairly small form, distinct from and probably not closely related to the gigantic American genera. It may be a relative of the African Tornieria; or an ancestral titanosaurid. During this time, Europe consisted of a number of islands; the small size of "C." greppini indicates that this could be an island-dwelling pygmy species (like the dwarf elephants of Pleistocene Malta). The dating of this formation is uncertain, but is tentatively given as Tithonian. (MAK 000218).
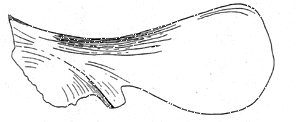 Rebbachisauridae: Agustinia?, Rayosaurus?
Rebbachisauridae: Agustinia?, Rayosaurus?
Range: lwK-upK of Afr, SAm, perhaps EEur.
Dental battery similar to hadrosaurs in anterior jaw; short basipterygoid processes (?); nares above orbit; quadratojugal greatly expanded in front of orbit; supratemporal, infratemporal fenestrae slit-like and nearly closed; pocket on posterior surface of quadrate; cervical ribs short but slender; neural spines long ("sail back"), not split; large pleurocoels on all dorsals; anterior caudals amphiplatyan; no forked chevrons; whiplash tail; distal ends of ischia elongated; forelimb/hindlimb 0.80; some with osteoderms with or without spikes (associated with the long neural spines).
Image: Right scapula of Rebbachisaurus gerasbae from Lavocat (1954). The actual length was 117 cm. This is the only figure in Lavocat's very brief description R. gerasbae, the type genus and species. Additional material of R. gerasbae may have been found in Morocco, but again there is nothing beyond a very short, preliminary report. Monbaron (1978). 010604.
Links: DinoData: Rayosaurus; sauropoda; Sauropodomorphs October 1); Notes on Rebbachisaurs and Tehuelchesaurus (longish); Rayosaurus; EVOLUTION OF HERBIVORY.
Note: much of the description here is based on Rayosaurus which is not securely placed in this group.
References: Lavocat 1954); Monbaron (1978); Sereno et al. (1999); Watterson 1989).
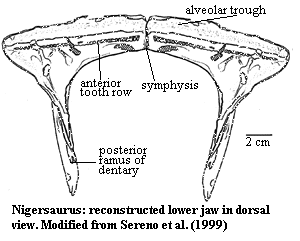 Nigersaurus:
Nigersaurus:
Range: mK of Africa.
<15m; teeth small, enamelled on labial side; anterior dental batteries with ~600 teeth; short, very wide & square rostrum (similar to Antarctosaurus, a possible Rebbachisaurid from SAm); $ tooth rows "oriented strictly transversely," forming straight, rake-like line across front of upper and lower jaws; $ tooth rows extend laterally beyond line of jaw; teeth with enamel much thicker on outer face, forming sharp cutting edge; ~7 replacement teeth for each position stacked in dentary & maxilla for total of 600 teeth; tooth row preceded by lip-like trough; lower jaw also extends outward to the sides somewhat like a duckbill; nares retracted as in advanced diplodocoids; $ elongated, narrow frontal with marked cerebral fossa; neural spines undivided; presacral vertebrae have simple, undivided neural spines with large, paired, sharp-rimmed pneumatic spaces (pleurocoels) under them on the dorsal vertebrae; broad, paddle-shaped scapulae as other rebbachisaurs; $ prominent rugosity on medial face of base of scapular blade.
Image: Using the best fit with the figure of Sereno et al. (1999), the tooth row was not quite transverse.
Note: Sereno et al. (1999) state that the unusual shape of the jaws and numerous teeth suggest Nigersaurus browsed on low-lying vegetation. They mention, but do not describe, a titanosaurid from the same location (Tegama group), as well as large hadrosauroids, and Ouranosaurus. 010605.
 Rebbachisaurus(=Limaysaurus?)
Rebbachisaurus(=Limaysaurus?)
~20m; teeth slightly curved; teeth spoon-shaped, and strongly se ; basipterygoid process oriented anterolaterally (not ventrolaterally); infratemporal fenestra extends to anterior margin of orbit; supratemporal fenestra faces laterally; cervical vertebrae with deep lateral excavation, divided into 2 by accessory lamina; very tall, undivided neural spines on dorsal vertebrae (presumed sailback -- even in the poorly known R. gerasbae, one of the neural spines is 145- 150 cm tall while centrum was ~20 cm long.) with deep pleurocoels in centra (two principle pleurocoels in R. gerasbae, separated by fairly small layer of bone); hyposphene- hypantrum articulations absent; substantial transverse processes; broad, paddle-like scapular blade, gastroliths.
Links: DinoData: Rebbachisaurus; Home Page; Paleontology and Geology Glossary: R; 0253333490.l.gif; GAIA Vol. 11, December 1995; SAMC NEWS 5; Musee dinosaures - sauropodes (French -- with image of the spine mentioned by Lavocat).
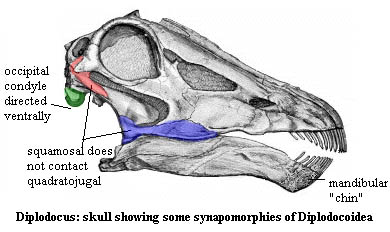 Diplodocoidea: Diplodocus + Dicraeosaurus.
Diplodocoidea: Diplodocus + Dicraeosaurus.
mJ-upK of Afr, NAm., Eur., Asia.
Characters: Skull long and low; teeth restricted to very front of mouth and pencil shaped; $? mandible forms thin, sharp "chin"; $ transverse narrowing of the rostral end of the premaxilla [U99]; premaxilla elongated posteriorly; $ elongation of the subnarial foramen [U99]; $ extreme retraction of the external nares [U99]; nares joined together at top of skull, above orbits, facing dorsally; narrow crown of skull; lateral temporal fenestrae narrow; $
squamosal does not contact dorsal process of quadratojugal; $? caudal surface of quadrate not excavated; $ occipital condyle directed ventrally; $? cervical vertebrae with concave ventral surfaces; $? cervical ribs very short (shorter than centra); $? bifurcated neural spines in (at least) posterior cervicals & anterior dorsal vertebrae; $ anterior caudal centra with slightly convex posterior surface (as opposed to deeper convexities in titanosaurs); $ expanded caudal ribs; most chevrons skid-like; whip-like tail; $ distal end of ischium dorsoventrally expanded; forelimbs much shorter than hindlimbs; loss of calcaneum from ankle.
References: Sereno 1999); Upchurch (1998); Upchurch 1999) [U99].
Image: See Diplodocomorpha.
Notes: Sereno (1999) asserts a mJ origin for this group. Short forelimbs said by some sources to implicate tripodal "rearing." -- but why should this be? Diplodocoidea, as used here, is equivalent to "Node U" of Upchurch (1998). A number of characters of the tail chevrons from Upchurch (1998) have been omitted as incomprehensible. 010907.
 Diplodocidae:
Diplodocidae:
40+ m long. Protruding snout; small, peg-shaped teeth confined to anterior jaw; $ tooth crowns lack grooves on outer face; $ ectopterygoid processes of pterygoid anterior to lacrimal, reduced and do not project ventrally; $ jugal forms substantial part of border of antorbital fenestra; $ rostral & dorsal processes of jugal form angle ~130°; quadrate rostroventrally inclined; external nares opening dorsally; $ distal ends of paroccipital processes rounded; very long necks; $ 15 cervical + 10 dorsal (+1 dorsosacral) vertebrae; tall neural spines; spines cleft; 80+ caudal vertebrae with "whiplash" of unornamented long bones at end; proximal caudals with moderately procoelous centra and wing-like transverse processes (fore-and-aft, sled-like chevrons); middle chevrons forked; distal ends of ischia expanded, meeting one another side by side; hindlimbs (much? a bit?) longer than forelimbs; $ calcaneum absent (but see Untitled Document); pes phalanx II-2 anteroposteriorly compressed; one specimen known with keratinous (?) spines; tripodal stance (2 legs + tail)?
References: Upchurch 1998).
Image: [1] An old, AMNH-style image of Diplodocus, almost as extinct as the sauropod -- but still interesting art and good anatomy. Unlike many newer illustrations, the proportions are correct, even if the posture and locale are unfashionable. [2] This taxon includes a number of other forms which are not yet dealt with in Palaeos. 010604.
Apatosaurinae: Apatosaurus
Range: Late Jurassic of North America
Phylogeny: Diplodocidae : Diplodocinae + *.
Inroduction: The apatosaurs constituted one of three distinct evolutionary lines of West Laurasian diplodocids; the other two being the barosaurs and the diplodocii. The apatosaurs are distinguished by their heavy build, relatively shorter but very thick neck, and lightly built forelimbs. Like all the diplodocids, the apatosaurs were huge terrestrial herbivores with a snake-like neck and whiplash tail, who probably fed on lower crown layers of trees and on undergrowth. Species include:
Apatosaurus excelsus (Marsh 1879)
syn. Brontosaurus, Elosaurus)
Morrison Formation; Wyoming, Utah, and Oklahoma, United States
Late Kimmeridgian to Early Tithonian
6 partial skeletons without skulls, hundreds of post-cranial elements.
Length: 18 to 21 metres; Weight: 13 to 20 tonnes
Apatosaurus ajax (Marsh 1879)
syn Atlantosaurus, Brontosaurus)
Morrison Formation; Colorado, United States
Early Tithonian
2 partial skeletons, braincases,
Length: 24 metres, Weight: upto 30 tonnes
The last and largest of all the brontosaurs, this was a huge animal, as big as the bigger brachiosaurids. The humerus or upper arm bone alone measured 2 metres in length, compared with 1.75 metres maximum for A. exelsus. It seems that, as the Morrison ecosystem progressed, there was a tendancy for these giants to evolve into larger and larger forms. The apatosaurs, diplodocines, camarosaurs, brachiosaurs, allosaurs, and stegosaurs all followed this trend, so the early Tithonian dinosaurs were among the most gigantic of these beasts that are known. The same tendancy to increased science also occurs at the very end of the Cretaceous (Campanian-Maastrichtian); in both cases this progession to gigantism was followed by a mass extinction. (MAK
000218)
Diplodocinae:
Range: Late Jurassic of Africa, North America
Phylogeny: Diplodocidae : Apatosaurinae + * : Barosaurus + Diplodocus. 020324.
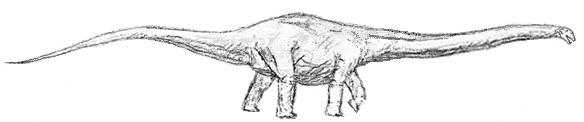 Barosaurus:
Barosaurus:
Range: Late Jurassic of Africa and North America
Phylogeny: Diplodocinae : Diplodocus + *.
Introduction:
 Barosaurus gracilus Janesch 1961
Barosaurus gracilus Janesch 1961
Middle Saurian Bed, Tendaguru Beds, Mtwara, Tanzania
[Middle or Late Kimmeridgian, Central Gondwana]
Many isolated limb elements; altogether the remains of at least 15 individuals
Overall length about 16 metres; live weight about 4.2 tonnes
B. gracilus is the earliest and smallest of the barosaurs, a line of sauropods which, like the mamemchisaurs, developed extraordinarily long necks. These enabled the beasts to both graze over a wide area of ground or swampland, and feed from the foliage of tree-ferns and cycads. Although the skull is not known, they probably had a delicate Diplodocus-like head, with weak peg-like teeth that could only have been useful for stripping leaves or on soft vegetation, and it is less likely that they fed much on the conifer forests that sustained the great brachiosaurs.
Occuring on both sides of the Jurassic Proto-Atlantic seaway, Barosarus may have evolved from an ancestral form some time during the late Callovian or early Oxfordian. In the moist Tendaguru rainforest megafauna there was only one evolutionary line of diplodocid, the small African barosaurs. But, in the drier American Morrison megafauna, are found three distinct lines: large barosaurs, Diplodocus, and the famous apatosaurs.
Initially, the barosaurs were a relatively unimportant element of the Tendaguru megafauna, making up 10% of identified specimens in the Middle Saurian Bed, and 4% of the actual megafauna biomass. Barosaurus is much more common in the succeeding Lower Transitional sands, where it accounts for
50% of identified specimens and 40% of the megafauna biomass. With the eventual dissapearance of the brachiosaurs in this region (possibly due to changing environmental factors) the baraosaurs increased in size and number, and became the dominant element of the Tendaguru fauna.
Barosaurus africanus (Fraas 1908)
syn. Gigantosaurus)
Upper Saurian Bed, Tendaguru, Mtwara, Tanzania
late Kimmeridgian/early Tithonian
More than 3 partial skeletons, a few skull elements, isolated postcrania; altogether the remains of at least 41 individuals
Barosaurus africanus: Size of selected skeletal elements
| Size in metres |
humerus
upper arm) |
ulna
lower arm) |
scapula
shoulder balde) |
femur
thigh bone) |
| average sized individual |
0.97 m |
0.74 m |
1.34 m |
1.34 m |
Overall length 20 metres;
Shoulder height about 3 metres; Hip height about 4 metres
Live weight 8.3 tonnes
Strangely, the great Brachiosaurus brancai, so common during the earlier Tendaguru, is totally absent in the later fauna. Instead, we find a larger species of Barosaurus, and a less abundant newcomer, Tornieria. In numbers Barosaurus africanus dominates the later Tendaguru fauna the way Brachiosaurus dominated the earlier fauna, being more common than Dicraeosaurus and Torniera put together. But in individual bulk Barosaurus is only half the weight of Brachiosaurus, and lived a different lifestyle. Whereas Brachiosaurus browsed giraffe-fashion from high branches, Barosaurus fed on undergrowth and low branches, its enormously elongated snaking neck conveying the head to select vegetation.
Barosaurus lentus Marsh 1890
syn. B. affinis)
Morrison Formation; South Dakota and Utah, United States
Early Tithonian
5 partial skeletons without skulls, isolated limb elements,
Overall length 23 to 27 metres; Weight about 15 tonnes
The American form is the largest of the three Barosaurus species; and identical to Diplodocus apart from its elongate neck. It has been suggested that it should be included in the latter genus. But the cervicals (neck vertebrae) are 33% longer, distinguishing Barosaurus as a distinct type. (MAK 000218).
Links: DinoData Dinosaurs B011 BAROSAURUS; Paleontology and Geology Glossary- Ba; BAROSAURUS; Barosaurus German); DinoNews.net - Le dinosaure Barosaurus French); Dino Land Travels Database Carnegie Museum- Barosaurus Bones; Barosaurus Display; Barosaurus Display.
030822.
 Diplodocus:
Diplodocus:
Range: Late Jurassic of North America
Phylogeny: Diplodocinae : Barosaurus + *.
Introduction: Diplodocus longus
Morrison Formation; Colorado and Utah, United States
Late Kimmeridgian
Length: 25 metres. Weight: 10 tonnes
The earliest American Diplodocus, a lineage of huge slender sauropods, it gave rise to D. carnegii, which in turn was supplanted by the even bigger D. ("Amphicoelus") altus. Bakker 1986).
Diplodocus carnegii Hatcher 1901
Morrison Formation; Wyoming and Utah, United States
Early Tithonian
5 skeletons without skulls, 2 skulls, hundreds of isolated post-cranial elements,
Length: 27 metres, Weight:10 tonnes
The successor and probable descendant of D. longus, Diplodocus carnegii is a slightly larger form. (MAK 000218)
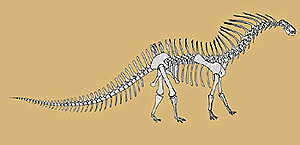 Dicraeosauridae: Amargasaurus, Dicraeosaurus. Small sauropods (some not much bigger than a horse)
Dicraeosauridae: Amargasaurus, Dicraeosaurus. Small sauropods (some not much bigger than a horse)
Introduction: Dicraeosaurus hansemanni Janensch 1914
Middle Saurian Bed, Tendaguru Beds, Mtwara, Tanzania
Skeleton lacking skull and forelimbs, 2 partial skeltons, altogether postcrania of about a dozen specimens
Overall length about 13.2 metres; weight 3.3 tonnes
Dicraeosaurus hansemanni: size of selected skeletal elements
| Size in metres |
humerus
upper arm) |
ulna
lower arm) |
scapula
shoulder balde) |
femur
thigh bone) |
| average sized individual |
0.75 m |
0.498 m |
1.35 m |
1.21 m |
A relatively small sauropod, Dicraeosaurus or "forked lizard" gets its name from the high, forked spines jutting up from the vertebrae. Lambert (1983). These were used to anchor powerful neck and tail muscles. The neck was very short, consisting of only 11 or 12 vertebrae, and the head was - for a sauropod - unusually large in proportion to the body. The forelimbs were very short (humerus to femur ratio 62%) but stoutly built. This was obviously a low-feeder, grazing on the undergrowth and low trees such as cycads. It is also the most primitive of the Diplodocoidea, presumably a late survivor of a line that must have appeared in the late Bathonian, although no traces of these early Dicraeosaurids have been found.
Dicraeosaurus was a common element in the Tendaguru megafauna, making up 22% of identified specimens in the Middle Saurian Bed, but (because of its small size) only 7.5% of the actual megafauna biomass.
Dicraeosaurus sattleri Janensch 1914
Upper Saurian Bed, Tendaguru, Mtwara, Tanzania
late Kimmeridgian/Early Tithonian
2 partial skeltons without skulls; altogether postcrania of about a dozen specimens
Overall length about 8.5 metres; Weight 1.0 tonnes
D. sattleri is considerably smaller than the earlier D. hansemanni; with a weight of only a tonne, and a hip height of about 2 metres, it had a body about the size of a large draft horse, with a long tail at one end and a snaky neck at the other. We tend to think of sauropods evolving towards greater and greater size, as with the brachiosaurs and brontosaurs. But Dicraeosaurus illustrates the opposite trend, and shows that not all sauropods were huge. This was probably a lively animal, browsing on the undergrowth and the lower branches of the trees of the Tendaguru forest, and sometimes rearing up to reach higher morsels. It was always in danger from the carnivorous Ceratosaurus, and may have gained some protection by living in large herds, like wildebeast do today. (MAK 000218)
muzzle longer; $ frontals "coalesced" (fused?); nares fully retracted to top of skull; $ supratemporal fenestrae face laterally; $ postparietal fenestra present; $ "leaf-like" dorsolateral process from crista prootica (internal mouse ears?); long, subparallel basipterygoid processes; $ basipterygoid processes at ~20° angle; $ deep pit between bases of basipterygoid processes; jaw long & square; neck shorter than diplodocids (~12 vertebrae); $ pleurocoels lost in cervical vertebrae; cervical ribs short; long bifurcated spines along back from elongated (~4x height of centra) neural spines esp. Amargasaurus); dorsal vertebral pleurocoels reduced or absent; tail long & thin; forelimbs short; .
Note: are the long neural spines really convergent with rebbachisaurs, or is this the primitive condition for diplodocomorphs, convergently lost in some of the larger members of the group? (ATW 010606).
Image: Amargasaurus.

Unless otherwise noted,
the material on this page marked 'MAK' may be used under a
Creative Commons License.
{kind=link}