Stromatoporoidea - 2
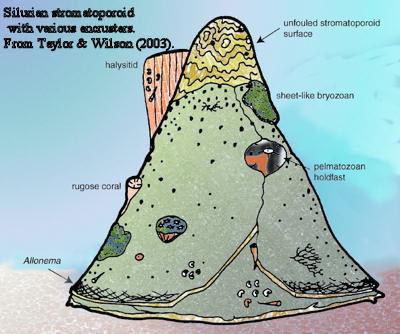 Growth Habit: the Calcite Empire. Like Romans or Englishmen, the stromatoporoids were colonial by nature, and often constructed edifices on a massive scale. These structures tend to be stratified, domal, or both. However (also like Romans or Englishmen), the very success of this monotonic system in gross depended on extraordinary adaptability and diversity in fine. Many stromatoporoids departed from the ground pattern and adopted other morphologies: bulbous, columnar, branching, or encrusting. Webby (2004); Benton & Harper (1997). And not just upper crust, either. One of the significant findings of recent work on reefs, ancient and modern, is the importance of cryptic communities -- organisms which lived under, within, or between the large calcifying elements. Wood (1998); Taylor & Wilson (2003). Stromatoporoids not only created these habitats, but sometimes lived in them as well.
Growth Habit: the Calcite Empire. Like Romans or Englishmen, the stromatoporoids were colonial by nature, and often constructed edifices on a massive scale. These structures tend to be stratified, domal, or both. However (also like Romans or Englishmen), the very success of this monotonic system in gross depended on extraordinary adaptability and diversity in fine. Many stromatoporoids departed from the ground pattern and adopted other morphologies: bulbous, columnar, branching, or encrusting. Webby (2004); Benton & Harper (1997). And not just upper crust, either. One of the significant findings of recent work on reefs, ancient and modern, is the importance of cryptic communities -- organisms which lived under, within, or between the large calcifying elements. Wood (1998); Taylor & Wilson (2003). Stromatoporoids not only created these habitats, but sometimes lived in them as well.
Associations: In Ordovician reefs, stromatoporoids are associated mainly with tabulate corals (and, less often, rugose corals), calcifying bacteria "algae") lingulid brachiopods, spirorbid worms, pelmatozoans, and bryozoans. Benton & Harper (1997); Taylor & Wilson (2003); Tapanila & Holmer (2006).
The diversity of organisms associated with stromatoporoids increased in the Silurian and Devonian, as the group spread beyond its Ordovician equatorial range. New inhabitants included goniatites, several new brachiopod groups, trilobites and crinoids. Tabulate corals remained important partners in building reefs, but rhodophytes (red algae) also became significant components of the complex. Stanley (2003); Wendt & Kaufmann (2006).
The Middle to Late Devonian was the zenith of reef-building -- not just for stromatoporoids, but probably for any biota, at any time in the history of our planet. Wood (1998). The stromatoporoids themselves produced a wide variety of types rarely seen in any other period: stromatoporoids which looked like mushrooms, or giant  cabbages, or bushes, or sandcastles -- well perhaps not sandcastles, but almost (see Wood, 1998: 192, fig 3, item 7). It would, in any case, be pointless to list their associations. Almost anything that lived in the sea at that time was living in or near a stromatoporoid reef, unless it avoided the continental shelves altogether.
cabbages, or bushes, or sandcastles -- well perhaps not sandcastles, but almost (see Wood, 1998: 192, fig 3, item 7). It would, in any case, be pointless to list their associations. Almost anything that lived in the sea at that time was living in or near a stromatoporoid reef, unless it avoided the continental shelves altogether.
One suspects that the main impetus for the evolution of enormous reefs in the Devonian may have been runaway competitive encrusting. Natural firm substrates were rare by the Ordovician, and became yet rarer. But see Taylor & Wilson (2003). The very success of the stromatoporoids and tabulate corals meant that they themselves gradually became the most widely available firm substrate in the oceans -- provided that an organism could encrust and outgrow the underlying sponge or coral, which was presumably not happy with the idea of becoming someone else's basement. Note that, the more that species A succeeded in encrusting and outgrowing species B, the greater the surface area it potentially provided to some third species C. That is, the more surface area B provided, the greater the selective reward to any other organism which could encrust and overgrow it.
 This may explain several features of Middle Paleozoic reefs which are otherwise difficult to understand. (1) Size. These reefs increased in size in a roughly exponential way from the Middle Ordovician to the end of the Devonian. This is what we might expect of a system in which competitive success in outgrowing others necessarily increased selective pressure to grow yet more. To put it another way, the more you succeeded in overgrowing the neighborhood, the greater selection pressure on others to specialize in overgrowing you. (2) Growth Rate. According to Wood (1998), Upper Devonian reefs grew about as fast as modern reefs, even without photosynthetic symbionts. This follows logically from the previous point. (3) Morphology. One puzzling aspect of stromatoporoids is why they kept building enormous skeletons even when, as in columnar forms, the living organism scarcely grew at all. This strategy may have allowed stromatoporoids to grow rapidly, to defeat encrusters, without expanding beyond the available space. (5) Cryptic Habitats. A number of writers have noted the extensive use of cryptic habitats: space under, between, or even within, other mineralizing organisms. This is sometimes said to be a response to predation, as in Recent reefs. However, the cryptic growth habit goes back to the earliest archaeocyath reefs -- before predation was likely to have been a significant problem -- and perhaps even further back. It may be better explained as an adaptation to the trophic equivalent of "Class C" office space.
This may explain several features of Middle Paleozoic reefs which are otherwise difficult to understand. (1) Size. These reefs increased in size in a roughly exponential way from the Middle Ordovician to the end of the Devonian. This is what we might expect of a system in which competitive success in outgrowing others necessarily increased selective pressure to grow yet more. To put it another way, the more you succeeded in overgrowing the neighborhood, the greater selection pressure on others to specialize in overgrowing you. (2) Growth Rate. According to Wood (1998), Upper Devonian reefs grew about as fast as modern reefs, even without photosynthetic symbionts. This follows logically from the previous point. (3) Morphology. One puzzling aspect of stromatoporoids is why they kept building enormous skeletons even when, as in columnar forms, the living organism scarcely grew at all. This strategy may have allowed stromatoporoids to grow rapidly, to defeat encrusters, without expanding beyond the available space. (5) Cryptic Habitats. A number of writers have noted the extensive use of cryptic habitats: space under, between, or even within, other mineralizing organisms. This is sometimes said to be a response to predation, as in Recent reefs. However, the cryptic growth habit goes back to the earliest archaeocyath reefs -- before predation was likely to have been a significant problem -- and perhaps even further back. It may be better explained as an adaptation to the trophic equivalent of "Class C" office space.
However, none of this is to be taken too seriously. For reasons detailed by Taylor and Wilson (2003), it is very difficult to get reliable data on interspecific competition among reef dwellers.
The Mesozoic stromatoporoids were less impressive, but still important in rebuilding the reef system after the end-Permian. Late Triassic stromatoporoids combined with sphinctozoan sponges and the newly important scleractinian corals. Stromatoporoids like Burgundia, Dehornella, and Acfinosfromaria created reef communities with red algae and scleractinians along high energy northern borders of the Tethys and on both shores of the Atlantic Ocean, as it began to open in the Western Tethys realm.
 So, are Stromatoporoids a Clade? Stromatoporoids were historically grouped with the Archaeocyatha (Rowland, 2001), and later with the demosponges (Leinfelder et al., 1996). Happily, we can agree with both contentions. This might be the consensus view, i.e., placing both Stromatoporoids and archaeocyaths on the demosponge stem. However, there is considerable uncertainty about who ought to be included among the "true" stromatoporoids for this purpose. The earlier Paleozoic forms seem very much alike, and are assumed to be monophyletic. Webby (2004). Similarly, it is hard to doubt the affinities of at least some Recent stromatoporoidal demospoges with Mesozoic stromatoporoids assigned to the same genus. Reitner & Engeser (1987) (Acanthochaetetes). However, it is still unclear (a) whether the Mesozoic Stromatoporoidea are a natural group and b) whether any of them are related to the Paleozoic bunch.
So, are Stromatoporoids a Clade? Stromatoporoids were historically grouped with the Archaeocyatha (Rowland, 2001), and later with the demosponges (Leinfelder et al., 1996). Happily, we can agree with both contentions. This might be the consensus view, i.e., placing both Stromatoporoids and archaeocyaths on the demosponge stem. However, there is considerable uncertainty about who ought to be included among the "true" stromatoporoids for this purpose. The earlier Paleozoic forms seem very much alike, and are assumed to be monophyletic. Webby (2004). Similarly, it is hard to doubt the affinities of at least some Recent stromatoporoidal demospoges with Mesozoic stromatoporoids assigned to the same genus. Reitner & Engeser (1987) (Acanthochaetetes). However, it is still unclear (a) whether the Mesozoic Stromatoporoidea are a natural group and b) whether any of them are related to the Paleozoic bunch.
Unnecessary confusion has been added to this area by a tendency to overplay the molecular results of Chombard et al. (1997). The paper is frequently cited for the proposition that Stromatoporoidea is polyphyletic, with some stromatoporoids being demosponges, and others Calcarea. Worse, it is said to support the idea that stromatoporoids are merely a "grade of organization" without phylogenetic meaning. This time, we're not going to let that phrase pass without challenge.
First, this is a 1997 study. A great deal of methodological progress has been made since then. Chombard et al. used only 100-200 "informative" base pairs from the 5' end of the 28S "gene" of about a dozen species. This required throwing out a lot of data which wasn't parsimony-informative, or couldn't be aligned, and represents a rather small sample of organisms. Different primer combinations were used for different species, which tends to stack the deck. No genes were cloned. Rather, the tests involved direct cloning of PCR products -- risky when the underlying RNA has a complex secondary structure. The paper is vague on the extent of intraspecific sequence heterogeneity and the procedures used to resolve the discrepancies
[10]. But, most fundamentally, this just isn't enough data on enough animals. See, for example, the remarkable effects of expanding the data and species numbers on the phylogeny of hexacorals. Daly et al. 2003). We are not questioning the conclusion that massive calcareous sponges are polyphyletic and "Sclerospongiae" an invalid taxon. We wish only to emphasize that this has absolutely nothing to tell us about stromatoporoids.
 Despite having started out strong, we must finish weak. We lack even an amusing speculation to offer about either of the questions in the first paragraph of this section. Our sole purpose was to argue that these are still very much open questions. But now, rather than attempt to crawl laboriously out of this embarassing rhetorical pit in full view of the reader, exposed to his disdain and sarcastic shouts of encouragement, we will draw a curtain around thes matters while distracting the reader's attention with a brief discussion of Hladil (2007).
Despite having started out strong, we must finish weak. We lack even an amusing speculation to offer about either of the questions in the first paragraph of this section. Our sole purpose was to argue that these are still very much open questions. But now, rather than attempt to crawl laboriously out of this embarassing rhetorical pit in full view of the reader, exposed to his disdain and sarcastic shouts of encouragement, we will draw a curtain around thes matters while distracting the reader's attention with a brief discussion of Hladil (2007).
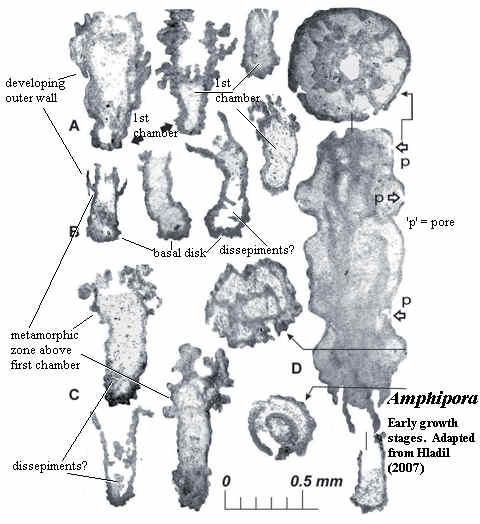
Hladil (2007) is not a "big" paper. It's a sort of "hmmm, this is interesting ..." kind of paper about the (mostly) Givetian-Frasnian stromatoporoid Amphipora. Amphipora is an usual stromatoporoid which is tall and thin, somewhat like the head of a horsetail, but sometimes branched. Hladil has been doing stratigraphy and geology in the Czech Republic for several decades and has seen a great many specimens.
In essence, his thesis is that Amphipora is rather archaeocyath-like in early development. One of his key figures is reproduced at right. (Our re-labeling is more aggressive in interpreting the structures than Hladil's original, as we must make the point more quickly). Amphipora develops from a single-walled "first chamber" with a basal disk. Above this chamber is a metamorphic zone, in which the wall delaminates, through a series of irregular pustules, some of which develop openings or pores. Eventually, there are two walls.
Hladil also emphasizes that neither the inner nor outer wall in the metamorphic zone seems to be continuous with the original wall of the first chamber. If so, this provides a broad developmental hint, not only about the origins of the Stromatoporoidea, but also about the relationships of some archaeocyath clades. That is, it suggests that the Monocyathida are truly basal, and that their wall may not be strictly homologous with either inner or outer wall of other archaeocyaths. Rather the separate walls derive from (as Hladil notes) an irregular series of growths resembling calcite bubbles, and suggestive of an originally secondary repair system. We are particularly enthusiastic about this observation, since it flanges nicely with our thrombolite speculations.
Phylogenetic History: We have covered a good bit of stromatoporoid history above. We repeat it, with a few extra details, in case you weren't listening the first time.
Some reviews have indicated that stromatoporoids are first known from the Tommotian Wood, 1998) or Botomian (Benton & Harper, 1997). However, Pseudostylodictyon from the Middle Ordovician of New York and New England may be the first "true" stromatoporoid. Id. Two distinct groups developed at that time, the Labechiida and the Clathrodictyida together with a few less well-known forms. The labechiids were particularly archaeocyath-like, with upwardly convex supporting plates. However, the water canal system is rarely preserved and is poorly understood. Webby (2004). By the end of the Middle Ordovician, large dome-like are found in North and South America. The labechiids were the dominant group in the Ordovician. They experienced two pulses of radiation in the Dariwillian and Sandbian, respectively. The Clathriodictyida also appeared in the Sandbian. Id.
The Labechiida declined in the later Katian and suffered further during the end-Ordovician glacial episode. The Clathriodictyida were the most successful group of stromatoporoids from that point until Famennian. Id. As discussed earlier, the Devonian was the high point of stromatoporoids. During this period, they formed vast reefs in association with red algae and calcifying bacteria (Wood, 1998; Stanley, 2003), sometimes hundreds of meters thick (Wendt & Kaufmann, 2006). This reef system was badly, but irregularly, disrupted during the Kelwasser event at the Frasnian-Famennian boundary. Benton & Harper (1997); Stanley (2003); Bond et al. 2004). During the Famennian, the Labechiids made a small comeback, but the Stromatoporoidea as a whole seem to have disappeared at the end of the Devonian.
Stromatoporoid-like animals reappear in the fossil record during the Late Triassic, forming reefs with scleractinian corals, spongiomorphs, hydrozoans and calcareous algae. Stanley (2003). During Anisian and Norian time, sphinctozoan and “stromatoporoid-like” sponges were at least as important as scleractinain corals. Id. These stromatoporoids re-established themselves as significant (but rarely dominant) parts of various reef-forming communities in the Late Jurassic. However, they were gradually replaced by rudist bivalves during the Cretaceous, and again disappear at the KT boundary. Id.; Benton & Harper (1997). In
1967, living stromatoporoid-like sponges were discovered in deep submarine caves and other out-of-the-way locations. Unfortunately, they are rare, scrupulously protected, and have thus remained rather poorly known.
It is still very uncertain whether stromatoporoids are a clade or simply a body plan on which various groups of sponges have converged, from time to time. Our prejudice is that they are probably a paraphyletic grade, but (properly constrained) not grossly polyphyletic. Most can probably be confined to a series of branches on the sponge stem group, a group which diverged at about the same time as the Archaeocyatha, and from about the same region of phylospace. Despite considerable stratigraphic and ecological work, we know very little about stromatoporoid anatomy, much less phylogeny. Like many sponges, stromatoporoids leave numerous and sometimes massive fossils, but the information content per gram of fossil material is perhaps the lowest of any major metazoan taxon. At the present time, we see only two notable rays of light: the rediscovery of living forms (analogues or ancestors, as the case may be), and the early results of Hladil's developmental work. Unfortunately, to quote Hladil, "[t]he examination methods are simple, but laborious." This is an important area for early metazoan phylogeny. It is grossly under-investigated, but not likely to yield big results from either improved technology or the discovery of some unique fossil specimen. ATW071008.
