Homoscleromorpha - 1
(= Homosclerophorida)
 Homoscleromorphas are a small clade of encrusting sponges. Homoscleromorphs or, sometimes, "homosclerophorids") are usually encrusting sponges, found all over the world in cool to tropical marine environments growing like underwater lichen on the surface of hard substrates. Ecologically, they usually favor relatively shallow marine shelves, but some species may also be found at depths approaching 1000 m. Muricy et al. 1998). They are anatomically simple, and often small and delicate, with few spicules -- or even none at all. Homoscleromorphs are said to be confusingly similar to the halisarcid demosponges. Fortunately, we ave no actual experience with the Halisarcida and are thus too ignorant to be deceived. In any case, you are unlikely to encounter a homoscleromorph, both because they usually grow in hidden (cryptic) places and because they are uncommon. Only a few genera are assigned to this group, the best-known being Oscarella and Plakina.
Homoscleromorphas are a small clade of encrusting sponges. Homoscleromorphs or, sometimes, "homosclerophorids") are usually encrusting sponges, found all over the world in cool to tropical marine environments growing like underwater lichen on the surface of hard substrates. Ecologically, they usually favor relatively shallow marine shelves, but some species may also be found at depths approaching 1000 m. Muricy et al. 1998). They are anatomically simple, and often small and delicate, with few spicules -- or even none at all. Homoscleromorphs are said to be confusingly similar to the halisarcid demosponges. Fortunately, we ave no actual experience with the Halisarcida and are thus too ignorant to be deceived. In any case, you are unlikely to encounter a homoscleromorph, both because they usually grow in hidden (cryptic) places and because they are uncommon. Only a few genera are assigned to this group, the best-known being Oscarella and Plakina.
They are historically important and may be the sister group of Eumetazoa. Although homoscleromorphs are an obscure and uncommon little group of sponges, they have sometimes played a pivotal role in the history of "spongology." Franz Schulze (who named the Plakinidae, the only homoscleromorph family) and Claude Lévi who named the Homoscleromorpha), both well-known sponge workers of circa 1870 and 1970 [16], respectively, both thought that the homoscleromorphs might be good approximations of the most primitive sponges. Leys & Ereskovsky (2006). However, in recent years,
 Homoscleromorpha has returned to center stage as a candidates for the living sponge taxon most closely related to the Eumetazoa, i.e., to all animals other than sponges. As discussed in tedious detail below and elsewhere, this suspicion is based on a number of characters which the homoscleromorphs share with higher animals, but not with other sponges. The theory is that sponges are paraphyletic. In other words, sponges are not just a side-branch of early animals. Rather, all animals actually evolved from among the sponges. According to this view, we aren't just related to sponges. We are sponges, merely highly evolved sponges.
Homoscleromorpha has returned to center stage as a candidates for the living sponge taxon most closely related to the Eumetazoa, i.e., to all animals other than sponges. As discussed in tedious detail below and elsewhere, this suspicion is based on a number of characters which the homoscleromorphs share with higher animals, but not with other sponges. The theory is that sponges are paraphyletic. In other words, sponges are not just a side-branch of early animals. Rather, all animals actually evolved from among the sponges. According to this view, we aren't just related to sponges. We are sponges, merely highly evolved sponges.
We have discussed some of the fine points of this theory, and a good deal of related stuff on biochemistry and embryology, in connection with the Demospongiae. The idea of sponge paraphyly has a lot of merit, and we have adopted it for purposes of the Palaeos cladogram. However, be aware that the concept of sponge paraphyly is still new and relatively untested. New ideas can only be tested slowly in sponge work, simply for lack of manpower. Here, we hope to take a somewhat less technical look at these potentially important animals, although we will inevitably slip into some discussion of our own peculiar (and probably wrong) ideas about sponge phylogeny.
Special Credit: special thanks to Dr. Ana Riesgo of the Centro de Estudios Avanzados de Blanes, Spain for help with sponge acrosomes and information on their phylogenetic distribution.
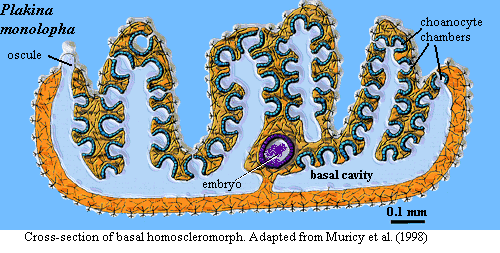 As usual, it is almost impossible to say anything useful about what a particular sponge group "looks like." The image of Plakina above is a good deal more typical than Oscarella. From the outside, the usual homoscleromorph sponge looks a bit like a dense collection of candle drippings. On the inside, the basic homoscleromorph body plan is ... rather basic. Its gross anatomy simply doesn't have any distinctive features. Like most encrusting sponges, homoscleromorphs have "top" and "bottom" surfaces which are quite different. The basal surface is flat and bears protruding spicules, specialized for attachment. The upper surface consists of a densely pleated/folded pinacoderm which supports flagellated pinacocytes. Embedded in the pinacoderm are numerous ovoid to spherical choanocyte feeding chambers. The choanocyte chambers may open into the external medium, into an internal cavity, or into both. Bergquist & Kelly (2004). Wastewater is directed into a basal cavity and out through an osculum.
As usual, it is almost impossible to say anything useful about what a particular sponge group "looks like." The image of Plakina above is a good deal more typical than Oscarella. From the outside, the usual homoscleromorph sponge looks a bit like a dense collection of candle drippings. On the inside, the basic homoscleromorph body plan is ... rather basic. Its gross anatomy simply doesn't have any distinctive features. Like most encrusting sponges, homoscleromorphs have "top" and "bottom" surfaces which are quite different. The basal surface is flat and bears protruding spicules, specialized for attachment. The upper surface consists of a densely pleated/folded pinacoderm which supports flagellated pinacocytes. Embedded in the pinacoderm are numerous ovoid to spherical choanocyte feeding chambers. The choanocyte chambers may open into the external medium, into an internal cavity, or into both. Bergquist & Kelly (2004). Wastewater is directed into a basal cavity and out through an osculum.
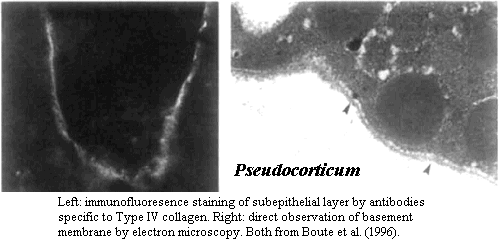 Eumetazoan-like basement membrane with type IV collagen. As we have said in innumerable other places, the homoscleromorphs are unique among sponges in having a basement membrane and, therefore, a true epithelium. We know this is a real basement membrane because it is composed largely of Type IV collagen, a protein not yet identified in any other group of sponges. Boute et al. 1996). We have extruded a large volume of words on the distinctive biochemistry of this stuff in a discussion of sponge skin. As if that were not enough, we have exhorted readers to examine Exposito's exposition on the subject (Exposito et al., 2002).
Eumetazoan-like basement membrane with type IV collagen. As we have said in innumerable other places, the homoscleromorphs are unique among sponges in having a basement membrane and, therefore, a true epithelium. We know this is a real basement membrane because it is composed largely of Type IV collagen, a protein not yet identified in any other group of sponges. Boute et al. 1996). We have extruded a large volume of words on the distinctive biochemistry of this stuff in a discussion of sponge skin. As if that were not enough, we have exhorted readers to examine Exposito's exposition on the subject (Exposito et al., 2002).
This requires significant changes in epidermal metabolism. The basement membrane in adult homoscleromorphs underlies the pinacoderm (outer surface) and the choanoderm (feeding chambers). Bergquist & Kelly (2004). As Maldonado (2004) points out, the existence of this barrier must have significant effects on sponge biology. In most sponges, cells of the pinacoderm are easily recruited from, and recycled to, the mesohyl. But homoscleromorphs have little mesohyl and the basement membrane makes migration between epidermis and mesohyl awkward. To the best of our knowledge, there is no substantial literature on the metabolism of the epidermis in homoscleromorphs, but it must necessarily be a more metabolically active and biochemically independent region than 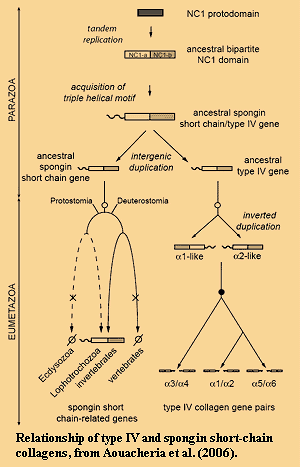 the outer skin of most sponges, i.e., more tissue-like and similar to the epidermis of eumetazoans. So, for example, Corticium develops tiny spicules which remain inside the cells of its epithelium, because the epithelial cells are separated from the internal mesohyl by the basement membrane. Maldonado & Riesgo (2007).
the outer skin of most sponges, i.e., more tissue-like and similar to the epidermis of eumetazoans. So, for example, Corticium develops tiny spicules which remain inside the cells of its epithelium, because the epithelial cells are separated from the internal mesohyl by the basement membrane. Maldonado & Riesgo (2007).
Some demosponges may also have a basement membrane. While a biochemically well-characterized basement membrane has only been identified in homoscleromorphs, morphologically similar structures have been described from at least one demosponge larva (Crambe crambe, see Maldonado, 2004), "and it is likely that other demosponge larvae also have this layer" Eerkes-Medrano & Leys, 2006). No such structure is known from calcareous sponges. Id.
Spongin is related to both type IV and short-chain eumetazoan collagens. The closest known relative of the basement membrane type IV collagen among non-homoscleromorph sponges is spongin, the characteristic collagen of demosponges. There are important structural relationships between spongin and Type IV collagen, as discussed in Exposito's review. Yet, judged by sequence alone, that relationship is fairly distant. We happen to think it peculiar -- in fact perverse -- to prefer sequence over structure when assessing the phylogenetic importance of a structural protein such as collagen. But only a tiny minority of biochemists seem to share our view, and it may be that we are mistaken in believing that this evidences a phylogenetic relationship between homoscleromorphs and demosponges. It is harder to dismiss the more recent work of Aouacheria et al. (2006). Aouacheria and co-workers (a group including Exposito) have shown that spongin is, in fact, not unique to demosponges. Short-chain spongin collagens are quite widely distributed among the Eumetazoa. However, nothing resembling either type IV collagen or spongin is known from the Calcarea. Thus, the natural conclusion is that homoscleromorphs, demosponges, and all non-sponge animals are more closely related to each other than any of them is to the Calcarea.
Homoscleromorphs also have many eumetazoan adhesion proteins. Another, perhaps related, suite of characters also unites homoscleromorphs with non-sponge animals. Homoscleromorphs have a nearly complete set of eumetazoan cell adhesion receptors and proteins which join the components of the cytoskeleton (microtubules, microfilaments, etc.). Nichols et al. (2006). Morphologically, homoscleromorphs have septate junctions between adjacent ciliated larval cells. Eerkes-Medrano & Leys, (2006). Again, these may also be present in some demosponges. Maldonado (2004). However, there is insufficient comparative data for this information to have much phylogenetic significance.
Possible artifacts related to sponge-associated organisms. We were so enthusiastic about the possibilities of tracing phylogeny through biochemicals that we spent several days plowing through unpublished dissertations in German, notably Blumenberg (2003). Since we speak no German whatsoever, this was a non-trivial exercise and came perilously close to doing actual work. 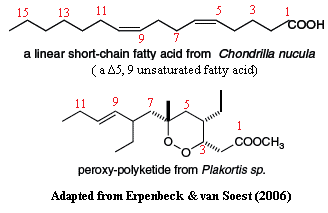 It might have been wiser to read Erpenbeck & van Soest (2006) (in English) before launching into this project. Those authors carefully explain the numerous pitfalls of "chemotaxonomy" -- pitfalls which are particularly deep when the technique is applied to sponges. Sponges are true animals, not just clonal aggregates; but they may also be viewed as cheap rooming houses for bacteria, protists, and fungi, along with other transients and undesirables. These boarders each bring their own metabolites and enzyme systems, and their population varies by sponge, by season, by reproductive state of the host sponge, or simply at random. So, chemotaxonomic statements have to be made with caution, and after broad sampling. As a practical matter, studies normally cannot be done with enough rigor to make indisputable statements.[17].
It might have been wiser to read Erpenbeck & van Soest (2006) (in English) before launching into this project. Those authors carefully explain the numerous pitfalls of "chemotaxonomy" -- pitfalls which are particularly deep when the technique is applied to sponges. Sponges are true animals, not just clonal aggregates; but they may also be viewed as cheap rooming houses for bacteria, protists, and fungi, along with other transients and undesirables. These boarders each bring their own metabolites and enzyme systems, and their population varies by sponge, by season, by reproductive state of the host sponge, or simply at random. So, chemotaxonomic statements have to be made with caution, and after broad sampling. As a practical matter, studies normally cannot be done with enough rigor to make indisputable statements.[17].
Excessive concern for autapomorphies. Another limitation of chemotaxonomy seems to be entirely self-imposed. As discussed elsewhere, traditional taxonomy looks for autapomorphies, while phylogenetics looks for synapomorphies. The former are unique characters which can be used to identify an organism. The latter are shared characters which can be used to identify relationships. One relevant example is a very odd family of compounds known as peroxypolyketides. Historically, these were regarded as unique to homoscleromorph sponges. Erpenbeck & van Soest, however, conclude that these compounds are now taxonomically useless. Why? Because, forsooth, peroxypolyketides have recently been isolated from three orders of demosponges, including the Haplosclerida -- sometimes suspected of sharing other traits with the Homoscleromorpha. Someone seems to have missed the point here. However, there are other reasons to be wary of relying on these particular compounds for phylogenetic purposes [18], so we will say no more.
 Demosponges and homoscleromorphs are united by Δ5,9 unsaturated fatty acids. In the last analysis, the only chemotaxonomic statement in which we place some tentative confidence is one we have mentioned elsewhere: Blumenberg's observation that Δ5,9 unsaturated fatty acids seem to be found in hexactinellids, demosponges, and homoscleromorphs, but not in calcareans. Blumenberg argues that this is significant, because the synthesis of Δ5,9 fatty acids requires a very specific enzyme system which is not known in the Calcarea. We like this result, but need to mention three problems, also discussed by Blumenberg. First, the substrate for the creation of medium- and long-chain Δ5,9 fatty acids is a short-chain saturated) bacterial fatty acid. Possibly, the Calcarea just don't associate with the correct bacteria, although this seems unlikely.Second, homoscleromorphs produce only a few, medium-chain (20-21C) varieties. They do not seem to synthesize the long-chain fatty acids made by demosponges and hexactinellids. Finally, the biochemistry of calcareous sponges is not exactly well-studied. Who knows what may be out there? Still, we count this as a modest point in favor of a demosponge-homoscleromorph sister-group relationship.
Demosponges and homoscleromorphs are united by Δ5,9 unsaturated fatty acids. In the last analysis, the only chemotaxonomic statement in which we place some tentative confidence is one we have mentioned elsewhere: Blumenberg's observation that Δ5,9 unsaturated fatty acids seem to be found in hexactinellids, demosponges, and homoscleromorphs, but not in calcareans. Blumenberg argues that this is significant, because the synthesis of Δ5,9 fatty acids requires a very specific enzyme system which is not known in the Calcarea. We like this result, but need to mention three problems, also discussed by Blumenberg. First, the substrate for the creation of medium- and long-chain Δ5,9 fatty acids is a short-chain saturated) bacterial fatty acid. Possibly, the Calcarea just don't associate with the correct bacteria, although this seems unlikely.Second, homoscleromorphs produce only a few, medium-chain (20-21C) varieties. They do not seem to synthesize the long-chain fatty acids made by demosponges and hexactinellids. Finally, the biochemistry of calcareous sponges is not exactly well-studied. Who knows what may be out there? Still, we count this as a modest point in favor of a demosponge-homoscleromorph sister-group relationship.
Also, possibly, by Δ6,X unsaturated fatty acids. In addition, but with less confidence, Blumenberg notes that some odd Δ6,X unsaturated fatty acids are found only in demosponges and Plakortis, a homoscleromorph. However, these may be entirely bacterial. Finally, he points a likely difference in the ability of sponge groups to synthesize sterols and probable marked differences in the sterols made. For example, aminosterols appear to be unique to homoscleromorphs. However, once again, comparative data from the Calcarea is insufficient to draw any conclusions. The literature on sponge biochemicals in general is sometimes obscure, but certainly large. At the same time, the coverage remains phylogenetically spotty.
CONTINUED ON NEXT PAGE
ATW080110
